经常有人抱怨:XXX免疫力太强了,免疫系统逮谁攻击谁,鸡蛋过敏/牛奶过敏/西红柿过敏,什么都不能吃。还有人甚至都有自免疫疾病(如甲减和类风湿性关节炎)了,免疫系统“杀疯了”,连身体正常细胞(或成分)都攻击。让人费解的是,这种“强悍”的免疫系统在面对呼吸道病毒时好像又“怂了”。这就是我们看到的过敏和自免疫攻击的人在流感季节也经常感冒/咳嗽,甚至感冒老好不了。为什么人的免疫系统会这样既强又弱,该弱的时候强,该强的时候弱呢?传统免疫学对免疫系统进行了精细划分,从免疫器官到免疫细胞到免疫活性成分,从先天免疫到后天免疫,从主动免疫到被动免疫,既零散又复杂,没有相关背景的人经常一看头都大了,完全看不下去。实际上,现代免疫学已经采取了更简单更好理解的划分。人体免疫系统主要包括两个相互独立的功能区:(统一)黏膜免疫系统和全身免疫系统,这两套免疫的工作方式和原理截然不同。本期食与心就带大家了解日常疑问,分析免疫问题到底出在哪里。为什么明明是在改善肠道,后来却不容易感冒了?过敏/自免疫时需要改善哪部分?感染时又应该强化哪部分?黏膜免疫系统,即统一黏膜免疫系统(Common / Unified Mucosal Immune System, CMIS / UMIS)。硬件包括:分布在呼吸道、消化道、泌尿生殖道等黏膜的淋巴组织,以及关腺体(如唾液腺、泪腺)。虽然主要位于黏膜部位,黏膜免疫系统统一的不是位置而是规则。- 统一的 诱导规则。抗原经黏膜进入→被 M 细胞 / 树突状细胞(DC )捕获→在黏膜淋巴组织中“低炎症方式呈递”,决定这是“黏膜型免疫”,而不是全身型。
- 统一的 迁移规则。在黏膜被激活的 B / T 细胞会表达特定整合素/趋化受体(如 α4β7、CCR9、CCR10),这些“地址码”决定它们是回到肠道 还是 迁移到鼻腔、肺、泌尿道等其他黏膜。这一步,才是“统一系统”的灵魂。
- 统一的 效应模式。黏膜免疫激活的最终输出高度一致,包括:分泌型 IgA(sIgA),局部耐受,屏障协作,而非强杀伤。
统一黏膜免疫系统不是“一堆黏膜免疫组织”,而是一个共享训练、共享地址、共享输出方式的免疫网络。在一个黏膜部位被“正确激活”的免疫细胞,可以被“定向迁移”到其他黏膜部位执行任务。在这个系统中,肠道是主教练,鼻腔和肺是前线,IgA 是共同语言。
在免疫球蛋白中,IgA(尤其是sIgA)是黏膜免疫的主体。IgA不激活补体系统(或仅弱激活替代途径),也不高效招募炎症细胞,对于病原体的主要策略是包裹、隔离、共存。因此IgA引起的免疫反应并不强烈,而是非炎症性、温和、局部化。
全身免疫系统(Systemic immune system)也称系统性免疫,主要涉及骨髓、脾脏、淋巴结和血液循环,即中枢免疫+外周免疫。全身免疫处理进入血液或组织的抗原,主要产生IgG、IgM等抗体,负责全身性防御和记忆响应。比如当感染突破黏膜屏障后,全身系统会激活细胞免疫(T细胞)和体液免疫。在全身免疫中发挥主要作用的抗体是IgG和IgM,IgM是初次应答主力(感染早期急升),IgG是次级/记忆应答主力(感染后期主导)。它们引发的免疫反应以炎症性、强效清除为主,与IgA的“温和预防”形成鲜明对比。IgG和IgM主要通过激活补体系统、调理吞噬和抗体依赖的细胞毒性(ADCC)等机制,直接杀伤或清除病原体,适合应对已入侵体内的细菌、病毒或毒素。这种反应往往伴随明显炎症(如红肿热痛),是全身免疫的核心。如果说全身免疫是“军队”,统一黏膜免疫系统就是“边防 + 外交 + 基础设施”。军队赢的是战争,边防赢的是时间与稳定。肠道是人体最大的免疫器官,但肠道免疫对整体免疫的最大意义,不是提供更多“兵力”,而是决定整个免疫系统的“训练方式、阈值和秩序”。肠道相当于免疫系统的“军校 + 指挥部”,而不是“主战场兵营”。肠道最重要的作用是设定“全身免疫反应阈值”。肠道是人体最大的抗原训练场,每天接触大量食物抗原、共生微生物、微量毒素等,肠道必须决定 :“什么该反应?什么必须忽略?”。这个判断规则会 通过 DC、Treg/循环细胞因子/神经—免疫轴 等传递给全身免疫系统,结果有两种:肠道决定免疫反应的性格 偏激还是稳健。肠道免疫教育正常时,肠道免疫稳态,其他黏膜也易安静;肠道失衡时,过敏和自免疫攻击等问题都可能出现。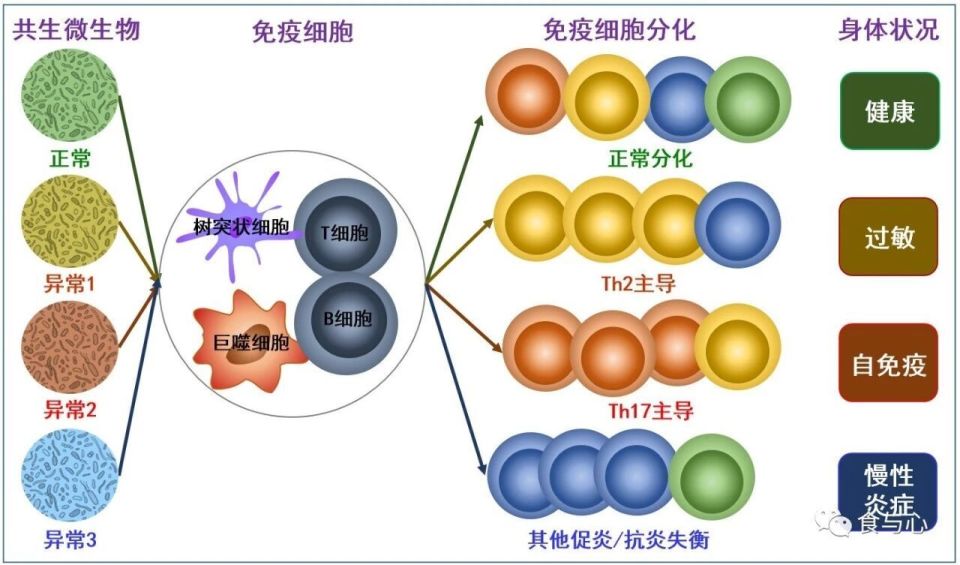 肠道通过“背景噪音”影响全身免疫效率。肠道稳态不好时:内毒素轻度入血,炎症因子长期偏高,全身免疫系统被噪音占用。这些会导致:感染时免疫反应慢、或者反应过度但效率低。肠道通过免疫微环境影响非黏膜器官。虽然肠道激活的免疫细胞很少直接驻扎在肝、脾、脑等非黏膜器官,但通过改变循环的细胞因子水平/肠菌代谢物(SCFA、色氨酸代谢物)/神经信号(迷走)等,肠道持续塑造这些器官的免疫“背景温度”。对于身体的整体免疫力来说,肠道更像“免疫理性中枢”。肠道免疫不是让全国多征兵,而是决定:什么时候该征兵,征什么样的兵,什么时候该停战。下面食与心将结合常见情况介绍,让你明白不同状况时哪些免疫系统在起作用。——以流感为例,了解黏膜免疫、全身免疫和肠道免疫的相互影响从免疫角度,流感不是“肺里单独发生的事”,而是一场从呼吸道黏膜发起、逐步动员全身免疫、最后再被重新收回到黏膜层面的系统性免疫事件。1. 第一阶段(0–24 小时):黏膜免疫“单独作战”气道会通过纤毛自动扶梯把黏液及其中被sIgA包裹/吸附的病毒送入咽部然后交给胃酸杀毒,或者通过鼻涕/痰排到体外 ,这就是气道特有的纤毛清除功能。不吸烟不饮酒和坚持健康餐饮,以及经常锻炼身体的人,纤毛清除功能够强,在这一步病毒负荷就能被降低,即便接触了病毒也只是无症状或轻症感染。此时,鼻腔 / 咽部 / 气道黏膜的先天免疫已启动,目标是局部解决,尽量不惊动全身免疫。上皮细胞感知到病毒RNA,分泌 I 型干扰素(IFN-α/β)和抗病毒肽等来对抗病毒。2. 第二阶段(1–3 天):黏膜 → 全身的“求援信号”病毒抗原被送往淋巴系统。树突状细胞(DC)将病毒抗原带到 鼻咽相关淋巴组织(NALT)和/或 支气管相关淋巴组织(BALT)。
肠道通过“背景噪音”影响全身免疫效率。肠道稳态不好时:内毒素轻度入血,炎症因子长期偏高,全身免疫系统被噪音占用。这些会导致:感染时免疫反应慢、或者反应过度但效率低。肠道通过免疫微环境影响非黏膜器官。虽然肠道激活的免疫细胞很少直接驻扎在肝、脾、脑等非黏膜器官,但通过改变循环的细胞因子水平/肠菌代谢物(SCFA、色氨酸代谢物)/神经信号(迷走)等,肠道持续塑造这些器官的免疫“背景温度”。对于身体的整体免疫力来说,肠道更像“免疫理性中枢”。肠道免疫不是让全国多征兵,而是决定:什么时候该征兵,征什么样的兵,什么时候该停战。下面食与心将结合常见情况介绍,让你明白不同状况时哪些免疫系统在起作用。——以流感为例,了解黏膜免疫、全身免疫和肠道免疫的相互影响从免疫角度,流感不是“肺里单独发生的事”,而是一场从呼吸道黏膜发起、逐步动员全身免疫、最后再被重新收回到黏膜层面的系统性免疫事件。1. 第一阶段(0–24 小时):黏膜免疫“单独作战”气道会通过纤毛自动扶梯把黏液及其中被sIgA包裹/吸附的病毒送入咽部然后交给胃酸杀毒,或者通过鼻涕/痰排到体外 ,这就是气道特有的纤毛清除功能。不吸烟不饮酒和坚持健康餐饮,以及经常锻炼身体的人,纤毛清除功能够强,在这一步病毒负荷就能被降低,即便接触了病毒也只是无症状或轻症感染。此时,鼻腔 / 咽部 / 气道黏膜的先天免疫已启动,目标是局部解决,尽量不惊动全身免疫。上皮细胞感知到病毒RNA,分泌 I 型干扰素(IFN-α/β)和抗病毒肽等来对抗病毒。2. 第二阶段(1–3 天):黏膜 → 全身的“求援信号”病毒抗原被送往淋巴系统。树突状细胞(DC)将病毒抗原带到 鼻咽相关淋巴组织(NALT)和/或 支气管相关淋巴组织(BALT)。
在淋巴结里,全身免疫被“定向动员”。初始CD4⁺ T 细胞分化为抗病毒的Th1(抗细胞内病原(如病毒、结核菌),促进巨噬细胞激活)和Tfh(滤泡辅助T细胞,能帮B细胞合成抗体)。B 细胞开始产生抗病毒的 IgM → IgG。需要注意的是:这一步不是“全面开火”,而是“为黏膜定制的全身响应”。3. 第三阶段(3–7 天):全身免疫反过来“支援黏膜”血液中的免疫细胞“回流”黏膜,表达黏膜归巢分子(α4β1、CCR5 等)的免疫细胞回到呼吸道上皮或肺泡间质。IgG 通过“非常规方式”进入黏膜。虽然 IgA 是黏膜主力,但IgG 可通过 FcRn 转运在严重感染中补位。这也是为什么:重症流感时,全身免疫反应强烈。感冒时的症状来自哪里?发热、乏力、肌痛主要是全身免疫信号(IL-1、IL-6、TNF-α)引起;而流涕、咳嗽则主要是黏膜免疫反应。4. 第四阶段(恢复期):全身免疫“撤兵”,黏膜免疫“收尾”病毒清除后,炎症必须被“主动关掉”。Treg ↑,IL-10↑、TGF-β ↑→黏膜上皮修复→纤毛功能恢复。如果这一步失败,或者不正确的药物投放,就会容易出现激活原本身体中的看似中性的细菌和真菌,导致继发细菌/真菌感染,结果发展为感染后咳嗽或鼻炎/哮喘加重的慢性炎症。免疫记忆的分配。黏膜免疫记忆主要由组织驻留记忆 T 细胞(Trm)和局部 IgA 记忆 B 细胞负责;全身免疫记忆则主要由循环 IgG和中央记忆 T 细胞负责。等下次再遇到病毒时,黏膜免疫先出手,全身免疫可能根本不需要动。真正强大的免疫系统,不是在病毒感染后启动更大的免疫反应,而是能把问题尽量解决在黏膜层面(不扩大成全身问题),在清除病毒后能迅速撤兵,恢复重建。如果黏膜免疫一开始就很弱,病毒会迅速复制→大量抗原进入血液→全身免疫被迫过度启动→炎症风暴风险增加。重症流感 ≈ 黏膜免疫失败 + 全身免疫被迫接管。很多人发现:感冒的时候,吃益生菌/酸奶确实好得极快,益生菌不是强化肠道免疫吗,怎么改善的呼吸道和全身免疫呢?实际上,呼吸道感染时,口服益生菌迅速改善,并非是“杀病原体”,而是通过“肠道 → 统一黏膜免疫系统”,把“更合适的免疫模式”送到了呼吸道。在接触益生菌后,肠道里的M 细胞、树突状细胞(DC)等会训练一批免疫细胞,比如IgA⁺ B 细胞和Treg / Th17 平衡型 T 细胞。这些细胞随后表达黏膜归巢受体(如 α4β7、CCR9 / CCR10),迁移到其他黏膜部位(比如鼻腔、咽喉、支气管)。这些免疫细胞不是“去杀病毒”,而是:提高局部 IgA,提高抗病毒反应的“精准度”,降低误伤。益生菌给免疫系统的信号是:“这里有微生物,但不需要全面开战。”造成的结果是抗病毒的I 型干扰素反应增强,炎症因子不过度升高。这能让呼吸道在面对病毒时:既能启动抗病毒程序,又不至于炎症失控。这正是绝大部分呼吸道感染者需要的状态。好的肠道微生物能产生短链脂肪酸(SCFA)和吲哚乙酸、吲哚丙酸等活性物质。这些物质会提高 Treg 稳定性,降低免疫系统“乱出手”的概率。所以补充益生菌后,肺部病毒清除效率即便没有更快,但肯定症状更轻、恢复更快。肠道是黏膜免疫的“中央训练营 + 调度中心”。感染时补充益生菌,相当于肠道益生菌给黏膜免疫系统一个“正确作战模板”,再由统一黏膜免疫系统把这个模板复制-粘贴到呼吸道,让抗病毒战争“打得更专一、更干净,而不是打得更狠”。这也是为什么:补充益生菌,能减少感染次数,缩短病程,减少感染天数,但对病毒载量下降速度影响不大。因为益生菌主要改善的是“宿主反应”,而不是“病原体本身”。所以有时候即便你还能从身体里检测出病毒感染阳性,但已经是无症状感染的感觉了。绝大多数过敏(尤其鼻炎、哮喘、结膜炎),问题主要出在“局部黏膜免疫失衡”,而不是“全身免疫系统坏了”。长期、反复、重度过敏 → 会逐步“牵连”全身免疫调控,但此时全身免疫属于“被拖下水”,不是起点。过敏,本质上是:“该容纳的没容纳,该快速清除的没清掉”。过敏性鼻炎时,鼻黏膜上皮屏障通透性 ↑,抗原更容易穿透,树突状细胞错误“教学”,这些会导致:T 细胞偏向 Th2 分化,B 细胞产生特异性 IgE,肥大细胞、嗜酸粒细胞高度待命。这是一套“局部自我放大的免疫回路”,不需要全身免疫异常就能发生。在过敏早期,全身免疫正常。但在长期 / 重度 / 多系统过敏情况下:长期 IgE 偏高,Th2 偏向开始“外溢”到外周,嗜酸粒细胞系统性升高,调节性 T 细胞(Treg)功能下降,慢性炎症因子背景升高。这时全身免疫“调控能力”开始出问题了,但并非免疫崩溃,而是“失衡”。因此,过敏不是“全身免疫有病”,而是“黏膜免疫这个前线岗位,长期在错误地拉警报”。类似于厨房装了个探测阈值太低的烟雾报警器,一做饭就报警。这里需要了解下IgE 。IgE是正常情况下五种免疫球蛋白中浓度最低的一种,主要针对寄生虫感染(生理保护作用)和过敏原(病理作用,如花粉、尘螨、食物、药物),特征是肥大细胞和嗜碱性粒细胞的脱颗粒,释放大量炎症介质,导致局部或全身急性炎症。
IgE引发的免疫反应最为剧烈和快速,属于I型超敏反应(也称速发型超敏反应),是“宁可错杀,不能放过”的模式。同样在黏膜表面,为什么免疫系统没有选择“温和、保护性的 IgA 路线”,而是走上了“激烈、放大反应的 IgE 路线”?这并非 B 细胞自己决定“不合成IgA合成IgE ”,而是黏膜树突细胞 + 局部环境给出的“危险判断”。IgA 是“和平共处”的抗体,IgE 是“紧急动员”的抗体。过敏的本质,是黏膜免疫在反复损伤中,被迫把日常环境误判为战场。IgA 路线的前提是:黏膜屏障完整+抗原重复出现但无损伤+低炎症背景。一旦环境“嘈杂”,免疫系统就会放弃耐受方案。
IgE 触发条件往往是:上皮破损(PM2.5、病毒、干燥),高氧化应激,神经肽释放(SP、CGRP),上皮释放 “危险信号”(如TSLP、IL-25、IL-33)。这些信号会告诉免疫系统:“狼来啦!这里不安全,可能有危险生物!”。此时免疫系统不再相信“这是一个可以单纯用 IgA 解决的环境”。IgE 有强大的“记忆放大器”:IgE 与肥大细胞 FcεRI 结合的半衰期长(细胞表面),再次遇到抗原 → 秒级反应;每一次发作,都在强化这条回路。IgE 反应本身会继续破坏黏膜:释放组胺 → 血管扩张、渗出;蛋白酶 → 上皮损伤;神经敏化 → 阈值下降。结果是:“黏膜越破 → 越不像一个适合 IgA 的环境”。这是过敏顽固的关键,一旦走上 IgE 路线,就很难回头。对于过敏性鼻炎,在肠道、神经节律和口腔正常的情况下,恢复鼻腔黏膜免疫,才能真正解决鼻腔问题。(解毒排毒第一关:你的鼻子挡住毒素了吗)过敏人群的感冒频率其实并不比一般人高,他/她们的全身抗病毒能力一般正常。问题在于“局部清除慢”,不是杀不死病毒,因为Th2 偏向(过敏)并不会完全抑制 Th1 抗病毒,但会让“炎症关停”更慢。所以常见现象是:病毒已清除,症状还在。容易过敏的人群流感时更容易出现 症状拖久/咳嗽、鼻炎反复/哮喘加重问题。——自免疫本质:黏膜免疫“点火”+全身免疫“烧成火灾”多数自身免疫病的起点,往往发生在“黏膜免疫—屏障—耐受”这一层,但真正决定疾病命运的,是后续“全身免疫刹车系统”有没有一起失效。
过敏常常可以起源于任何一个黏膜或皮肤屏障(鼻、肺、皮肤);自免疫疾病的“起火点”可以很多(比如鼻、皮肤、肺、肠),而“持续供油点”往往是肠道,是否烧成全身病则由全身免疫决定。
所以在过敏疾病干预时,即使只修复局部黏膜,也能看到改善;但自免疫疾病干预则完全不能绕开肠道,即使症状在皮肤/关节/甲状腺等非肠道器官。下面食与心以强直性脊柱炎为例,来介绍下疾病中的免疫变化。强直性脊柱炎(AS)尽管强烈依赖遗传,但“发病”和“进展”高度依赖黏膜免疫(尤其是肠道)与全身免疫的持续互动;没有黏膜触发,遗传只是“配了枪”,但不一定会开火。HLA-B27 是AS的“风险放大器”,不是病因本身。HLA-B27 阳性人群中,只有约 5–10% 最终发展为 AS;而在AS中,有10-20%是HLA-B27 阴性。HLA-B27 真正的问题不是“它存在”,而是它参与:- 错误折叠(misfolding) → 细胞内应激 → 炎症信号;
- 与特定肠道菌群(比如肺炎克雷伯氏菌)强烈相互作用;
AS 的“免疫火种”,极常来自肠道,而不是脊柱本身。虽然强直性脊柱炎的主要症状在脊柱,但其不是“脊柱的病”,而是“黏膜免疫失控后,选中了脊柱作为战场”。临床调查发现:60–70% AS 患者存在“无症状肠道炎症”,即使没有腹泻/腹痛/明显炎症。大部分患者有肠漏问题,微生物成分进入黏膜内,激活先天免疫。AS 患者常见菌群特征:Prevotella ↑,克雷伯氏菌增加(AS强相关),产短链脂肪酸(SCFA)的菌 减少。菌群紊乱的后果:诱导 Th17 / IL-23 轴 偏移,抑制了 Treg 的“刹车系统”,进入自免疫“攻击模式”。正常肠道应该教会免疫系统:“这些是共生菌,不要打”。但在 AS 中:IgA 异常,抗原呈递偏向炎症性,Th17 被过度扩增;这些导致免疫系统学会了“错误的敌我识别”。一旦黏膜免疫失衡,肠道产生IL-23进入循环→激活循环中的 Th17→Th17 表达 CCR6、α4β7 等迁移分子→Th17迁移到骶髂关节或脊柱附着点,启动自免疫攻击。全身免疫的核心轴线,如IL-23 / IL-17,TNF-α,和GM-CSF进一步加剧免疫反应。这也是为什么:抗 TNF、抗 IL-17对 AS 治疗效果显著。需要注意的是:这些药物“灭火”,但并不总能阻止复燃。原因就是:肠道黏膜源头还在持续供火。强直性脊柱炎的长期管理,不是“找一个最强的干预”,而是:用药物压住主轴,用运动重塑局部免疫,用节律和黏膜减少复燃概率。这是AS 管理的“地基”,不做=病程几乎一定推进。- 使用抗炎药物(以生物制剂 / 必要时 NSAIDs 为核心)降低结构进展风险、延缓脊柱融合。因为 AS 的核心病理是:IL-23 / IL-17 / TNF 驱动的附着点免疫炎症,不压住炎症,问题会越来越严重。
- 规律、长期、结构化的“免疫友好型运动”。运动在这里相当于慢变量免疫治疗,不是“锻炼身体”,而是:持续给附着点一个 “这是正常负荷,不是损伤信号”的免疫输入。不做这件事≈ 炎症信号长期存在,即使药物有效,也更容易反跳。运动在这里相当于慢变量免疫治疗。
睡眠与节律稳定(被严重低估)。睡眠是“每天一次的抗炎治疗”。AS 患者的共同点:夜间炎症活跃,清晨最重。而节律紊乱 → IL-17 基线升高,睡眠差 → 炎症关停失败。轻度AS患者药物可降至第二层级,但运动和睡眠节律仍是必需干预策略。做得好 = 药效更稳、复发更少。这一层决定:“你是在被动压病,还是在主动减少复燃概率”。黏膜免疫稳定(尤其是肠道),这是 AS 的“源头层”,但不是急救层。有意义的前提是:不折腾、不激进、长期一致。真正有价值的目标只有三个:肠屏障不再反复破,肠道炎症背景降低,Th17 的“供给源”减少。这不是“治好 AS”,而是 减少免疫火源。感染控制与疫苗策略(精准,而非泛化)。使用免疫抑制药物会降低抗病毒能力,因此用药期间流感重症/继发肺炎风险增加→ 是 AS 炎症波动的重要触发器,注意此时疫苗是“防并发”,不是“治 AS”。神经系统调控(降低压力、激活迷走神经),能降低炎症放大,缩短恢复期,减少疼痛感知,这是“放大器调小”, 不是“关掉病因”。饮食结构调整(而非极端方案)。避免反复诱发肠道炎症,维持体重与代谢稳定,注意 长期一致 > 短期极端。迷信“增强免疫”,比如各种免疫刺激补剂,高强度训练硬顶。对 AS 来说:完全拒绝药物,单靠生活方式。这不是“自然疗法”,而是:把一个结构性免疫病,当成了功能失调。不管是感染、过敏还是自免疫问题,康复过程都绕不开的一个步骤就是睡眠。为什么睡眠这么重要?神经系统能通过「迷走神经 + 交感神经 + 神经肽 + 中枢节律」多通路、分层地塑造黏膜免疫和全身免疫的“基线状态”。迷走神经是“免疫教学的纠偏器”,作用不是“关掉免疫”,而是恢复耐受。迷走神经高张力本质是:副交感系统的“刹车能力强、反应快、恢复好”。迷走神经高张力能激活胆碱能抗炎反射,抑制“过度炎症”,但不抑制防御性免疫。关键机制:迷走神经 → 乙酰胆碱→抑制巨噬细胞 TNF-α / IL-1β / IL-6→降低炎症“放大”。在黏膜环境中,迷走神经高张力,可增强肠上皮紧密连接,促进杯状细胞黏液分泌,提高肠道 IgA 转运,这些会 减少抗原跨屏障进入 → 从源头降低免疫激活。交感神经是迷走神经的对立面。交感神经高张力本质是:交感系统的“启动阈值低、持续时间长、关停慢”。交感神经高张力会引起炎症放大偏置。短期交感激活,可动员免疫细胞,有利于应急防御。但长期交感高张力(慢性压力),会诱发β2-AR 信号失衡,Th17增加,Treg 减少,IgE 偏移。因此 慢性压力 = 炎症型免疫教学。交感神经重塑免疫“去向”。交感信号会改变免疫细胞趋化因子表达,影响它们“去哪儿”,这对强直性脊柱炎极其重要,此时Th17 更容易迁移到附着点,骶髂关节成为“免疫热点”。神经肽不是神经系统影响免疫的唯一语言,但在黏膜、附着点和炎症前线,它是最贴近战场、最能左右结果的语言。常见神经肽中,P物质促炎,CGRP双向偏保护,VIP强耐受。在黏膜和附着点,神经纤维可与免疫细胞直接接触。神经肽能即时改变:肥大细胞脱颗粒、DC 的抗原呈递方式、IgA / IgE 偏向、这是“毫米级”的精细免疫调控。- 昼夜节律会设定免疫阈值。夜间:Treg 活跃,组织修复优先;熬夜/失眠:炎症因子全天高位,自身免疫风险增加。
- 昼夜节律影响HPA 轴释放抗炎激素(主要是皮质醇)。皮质醇正常节律:免疫稳定;长期紊乱:免疫耐受下降,自身免疫风险上升。
睡眠本身就是一场神经-免疫的压力测试,睡眠质量是判断神经张力的“温度计”。
神经张力不是“开或关”,而是:系统在静息状态下,能否迅速、精准、可逆地被调动。迷走神经高张力不是“更放松”,而是“更能关掉不该持续的反应”。交感神经高张力不是“你太紧张”,而是“系统忘了怎么停下来”。用睡眠情况来推测神经张力,不能只看睡眠体验,需要看4个睡眠维度。正常情况下(迷走张力良好):入睡快(10–20 分钟),夜醒少,醒后能再睡,清晨自然醒,起床后恢复快。以下4种情况是夜间交感残留的经典表现,强烈提示交感高张力:- 夜醒(特别是凌晨 2–4 点醒),心率上来,再入睡困难。
- 耗竭型入睡。白天交感拉满,晚上直接“断电”,这不是健康迷走,是“累到关机”。
- 酒精 / 药物助眠。虽然入睡快,但深睡比例下降,夜间交感反跳。此时睡着 ≠ 修复。
- 炎症性嗜睡。睡得久,但醒来仍累,白天困倦。这是炎症在压系统,不是迷走张力高。
由于睡眠体验的主观性强,长期的睡眠趋势结合心率变异性(HRV)判断神经张力更准确。神经系统不是在“控制”免疫,而是在持续告诉免疫系统:“现在是安全环境,还是战时状态。” 迷走神经:告诉免疫“可以耐受”;交感神经:告诉免疫“要警惕”;神经肽:决定“局部怎么反应”;中枢节律:设定“反应阈值”。实际上,神经系统判断免疫系统【该修养】 还是 【该战斗】的主要标准就是睡眠。并不是躺床上就叫睡眠,放松的睡眠与迷走高张力相关,是免疫系统的耐受信号;而熬夜/失眠/辗转反侧则与交感高张力相关,是免疫系统的战斗信号。除了睡眠,增强迷走神经张力,让免疫系统明白【无需过度反应】的方法还有很多,比如大笑、哼歌、跟信任的人拥抱等。食与心温馨总结:人体免疫系统主要分为两个相互独立又相互依赖的功能分区:黏膜免疫和全身免疫。黏膜免疫的硬件是身体内外表面的黏膜免疫组织,肠道是其中最大组分。全身免疫则主要由循环中的免疫细胞和中枢免疫器官组成。黏膜免疫相当于边防和外交,全身免疫相当于军队和警察,而肠道则是最大的军校和指挥部。黏膜免疫的主要通用语言是IgA,引起非炎症局部免疫反应,相关症状比如感冒时的流鼻涕和咳嗽。而全身反应的主要通用语言是IgM和IgG,涉及猛烈全身性反应,相关症状比如感冒时发烧、肌痛、乏力。呼吸道病毒感染时,健康的免疫系统能通过纤毛清除功能和 I 型干扰素反应等将病毒拦截在黏膜部位,在病毒清除后也能迅速启动黏膜修复,感染者往往是无症状或轻症。而黏膜免疫能力低下时,病毒能迅速突破黏膜,长驱直入激活全身免疫,依靠猛烈的全身炎症反应消灭病毒,此时就会出现严重感冒症状;如果在病毒清除后,炎症反应仍没有停下,身体迟迟不能进入修复重建环节,出现各种后遗症。感冒时的很多严重症状往往不是病毒造成,而是炎症反应中伤及了无辜组织,所以病毒清除后有些症状仍会持续一段时间。补充益生菌,能通过稳定肠道免疫影响全身免疫,让抗病毒反应更加精准,减少殃及无辜,让免疫反应收尾更加干净,修复更快。过敏的本质是黏膜局部免疫耐受失败。当黏膜渗透性增加,上皮细胞反复接触抗原,树突状细胞根据局部环境判断“不安全,IgA解决不了问题”时,会促使B细胞合成IgE,之后黏膜免疫从温和/保护性的IgA反应转向激烈/自行放大的IgE反应。一旦再接触这种抗原,黏膜免疫就开始狂拉警报;暴露次数越多,免疫反应越激烈。
由于过敏的本质问题在黏膜免疫,修复时一定不能绕过黏膜的主体——肠道。无论是鼻腔、肺、皮肤还是眼睛过敏,干预时都需要先调好肠道免疫稳态,不然过敏症状暂时改善后也可能反复。如果说过敏主要是黏膜免疫问题,那自免疫更牵涉全身免疫。自免疫疾病的起火点可能在黏膜,但持续火上浇油点则在肠道,全身免疫的问题在于刹不住车,不能停止免疫反应。以强直性脊柱炎为例。肠漏后,免疫细胞在肠黏膜学会了错误的攻击模式——免疫向Th17 / IL-23 轴偏移,当这种错误的黏膜免疫信息进入循环,会激活循环Th17细胞,之后激活的Th17细胞会迁移到附着点(脊柱和骶髂关节),攻击脊柱和骶髂关节。因此,在强直性脊柱炎的治疗过程中,除了抑制脊柱的自免疫反应,还需要通过适当运动和睡眠节律强化全身免疫的“刹车能力”,当然也需要修复肠道屏障,减少自免疫攻击“火源”和肠道中的“火上浇油”。
需要强调的是:人体不管是排毒解毒,还是感染、过敏、自免疫,都需要充足的睡眠。因为睡眠是神经免疫调节的温度计,好的睡眠能让神经系统通知免疫系统 【可以耐受、可以停止炎症】,从而避免了过度炎症的问题。这方面的自然例证就是动物:家里的宠物病了之后自己知道要减少进食,多喝水,多睡觉。良好的睡眠一定是一种最省钱的康复之道。