好书推荐!《动物行为实验指南》电子版pdf,网盘发货
《动物行为实验指南》共674页,涵盖了常见的实验动物,如小鼠、大鼠和斑马鱼,详细描述了每一种行为测试的实验设计、测试设备、实验流程、评估指标、预期结果、常见问题及解决方法、数据分析、模型应用与局限性等各个方面。它通过快速引导,帮助研究人员高效地掌握实验的每个阶段,减少了查阅文献和寻找方法的时间,成为各类科研人员的重要参考资料。
《动物行为实验指南》共计收录了16种动物行为类型,包括焦虑抑郁、学习记忆、痛觉、运动、恐惧、社交、癫痫、操作、成瘾、视觉、痒觉、味觉、嗅觉、睡眠、斑马鱼行为以及常见动物模型等内容。每一类动物行为下,都详细介绍了多个经典的实验范式,涵盖了超过100种实验方法。
为确保其生存,动物必须能够对其环境中的威胁作出适应性反应。然而,支持灵活防御行为的精确神经回路机制仍不清楚。基于此,2025年1月31日法国国家科学研究中心Tihana Jovanic研究团队在Nature communications杂志发表了“Neural circuits underlying context dependent competition between defensive actions in Drosophila larvae”揭示了果蝇幼虫中的防御行为竞争背后的神经回路是复杂且动态的,这些回路能够根据环境线索的不同而灵活调整。通过在果蝇幼虫中使用神经元操控、基于机器学习的行为检测、电子显微镜连接组学以及钙成像技术绘制出了参与响应相互竞争的厌恶刺激时防御动作之间竞争的不同第二级中间神经元。研究发现机械感觉刺激抑制了逃避行为,转而倾向于惊跳行为,这是通过影响促进逃避行为的第二级中间神经元的活动实现的。激活这些神经元会抑制类似惊跳的行为,这表明惊跳和逃避行为之间的竞争发生在第二级中间神经元层面。最后,确定了一对促进惊跳行为的下行神经元,并且它们可能调节逃避序列,这些结果描绘了在感觉背景下被调节的惊跳与逃避竞争所涉及的路径。 图一 果蝇幼虫在面对机械刺激(如气流刺激)时逃避反应背后的神经回路作者首先研究了幼虫对气流刺激的行为反应是由一系列五个不同的、互斥的逃避动作组成的:拱起(Hunch)、弯曲(Bend)、后退(Back-up)、停止(Stop)和爬行(Crawl)。这些行为的基础是一系列由机械感受神经元、抑制性神经元以及投射神经元Basin-1和Basin-2构成的神经网络。进一步的研究表明,Basin-1和Basin-2的共激活导致了弯曲行为,而仅Basin-1的激活则引起了拱起行为。此外,A19c神经元被认为是通过接收来自Basin-2和Basin-4的输入来抑制拱起行为的。接下来,研究还发现了其他潜在的候选神经元,例如A08m和A08x,它们主要从Basin-1和Basin-3接收输入,并且在一定程度上也从Basin-2和Basin-4接收输入。该部分不仅揭示了参与果蝇幼虫逃避反应的神经回路的具体组成,还展示了如何通过调节不同神经元群体之间的相互作用来实现复杂的行为选择。 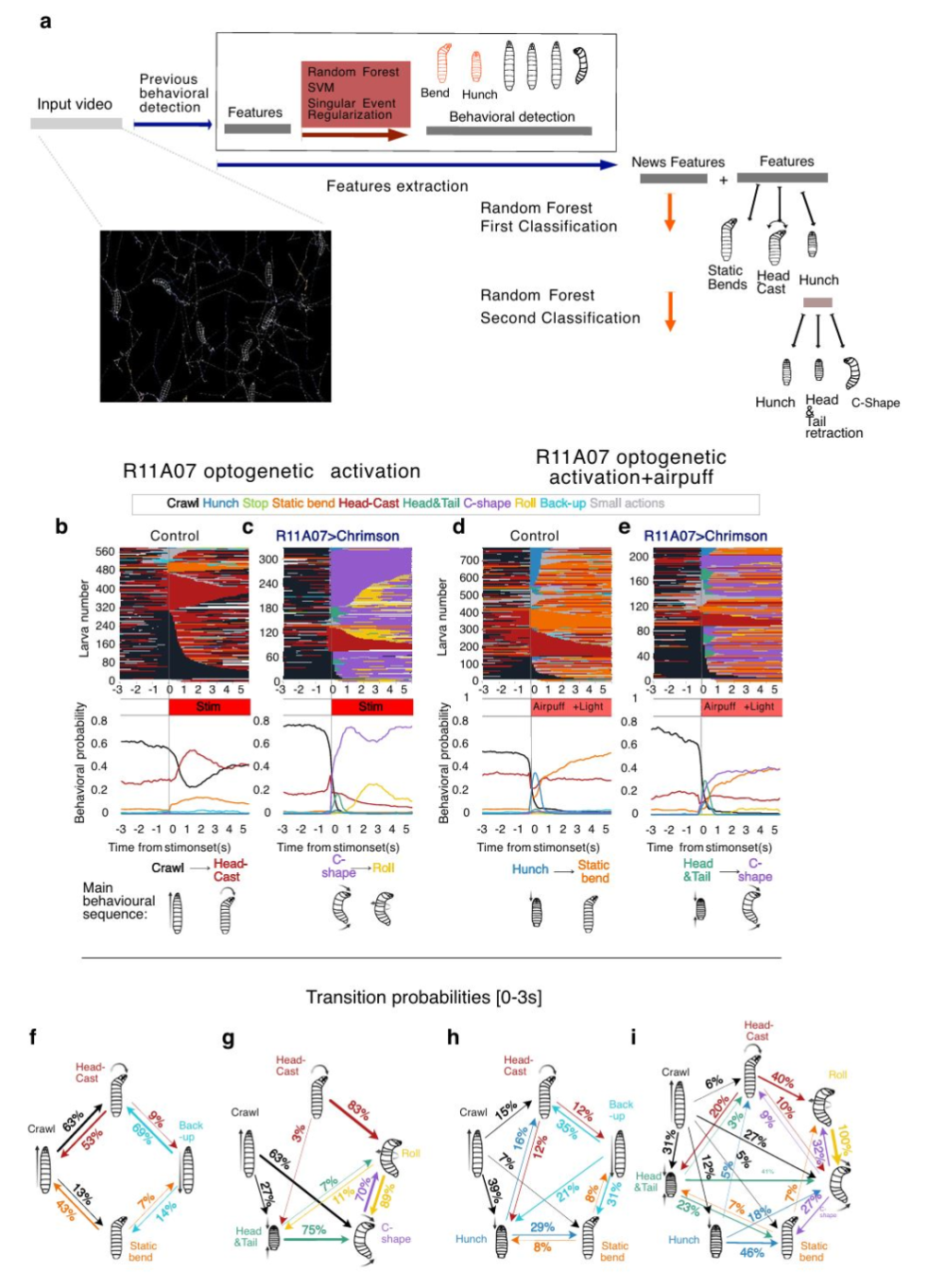 图二 利用自动化分类方法来分析果蝇幼虫在面对机械刺激时的行为反应
图二 利用自动化分类方法来分析果蝇幼虫在面对机械刺激时的行为反应
为了量化不同刺激条件下逃避序列中不同行为的概率,研究人员扩展了基于机器学习的行为分类算法,以检测头部-尾部收缩和C形弯曲动作。利用这些新的分类算法,研究者们分析了幼虫对气流刺激、单独的R11A07神经元光遗传学激活或两者结合的反应。结果显示,在仅接受光遗传学激活的情况下,大多数幼虫执行的是C形弯曲,随后通常是滚动行为。然而,当光遗传学激活与气流刺激结合时,幼虫除了执行上述逃避行动序列外,还会表现出拱起和静态弯曲(Static Bend)。在这些逃避行为(头部-尾部收缩、C形弯曲和滚动)中,相较于单纯的光遗传学激活,联合刺激导致了更多的头部-尾部收缩和较少的C形弯曲及滚动行为。这些结果揭示了不同的逃避序列是由R11A07>CsChrimson激活引起的,且取决于是否与机械刺激相结合。当激活与气流刺激结合时,头部-尾部收缩 > C形弯曲的序列更为明显;而单独的光遗传学激活后,则主要发生C形弯曲 > 滚动的序列。 图三 在R11A07品系中,胸部和腹部神经元具有不同的突触前连接性在R11A07品系中,胸部和腹部神经元具有不同的突触前连接性。R11A07驱动标记了两种类型的神经元:腹部的A19c和胸部的下降神经元(TDN)。TDN发出侧向下降投射,几乎跨越整个腹神经索(VNC,其神经系统的核心部分,位于身体的腹侧,从头部延伸到尾部)的长度。这种投射比腹部A19c轴突更为外侧和背侧,并且两种神经元都有轴突投射连接到对侧。这表明它们可能在控制需要不对称收缩的运动动作中发挥作用,如弯曲(Bends)、C形弯曲(C-shapes)和滚动(Rolls),TDN可能整合跨节段的机械感觉输入。通过分析该神经元所有突触前伙伴的分布情况,发现机械感觉输入占所有输入的15%。其他部分包括来自下降神经元的输入(31%),来自VNC本地神经元的输入(21%),以及来自上升神经元的输入(12%)。大部分TDN从脑和SEZ神经元接收到的下降输入是由轴突-轴突连接组成的,暗示高级神经元根据情境、状态或经验调节TDN的输出。相比之下,A19c主要从VNC内的本地和投射神经元接收输入(47%),其中Basins占了一半以上。只有8%的输入来自下降神经元,并且A19c不直接接收任何感觉神经元的输入。 研究结论揭示了在面对威胁时,惊跳(startle)和逃避(escape)这两种行为之间的竞争性互动是在二级中间神经元的水平上实现的,并且这种竞争不仅发生在早期处理阶段,即在选择惊跳型和逃避型反应之间整合内部状态信息的地方,还在更高级别的处理中得到了体现。这意味着在神经系统中对于适当防御行为的选择可能是分布式的,并可能整合有关内部状态和环境的各种信息。动物的防御反应不是单一、固定的行为模式,而是基于当前情境和个体状态的复杂动态过程。在不同的环境中,同样的动物可能会对相同的威胁做出不同的反应,这取决于它所接收到的感觉信息以及它的内部状态,如饥饿程度、疲劳状况或之前的经验等。

