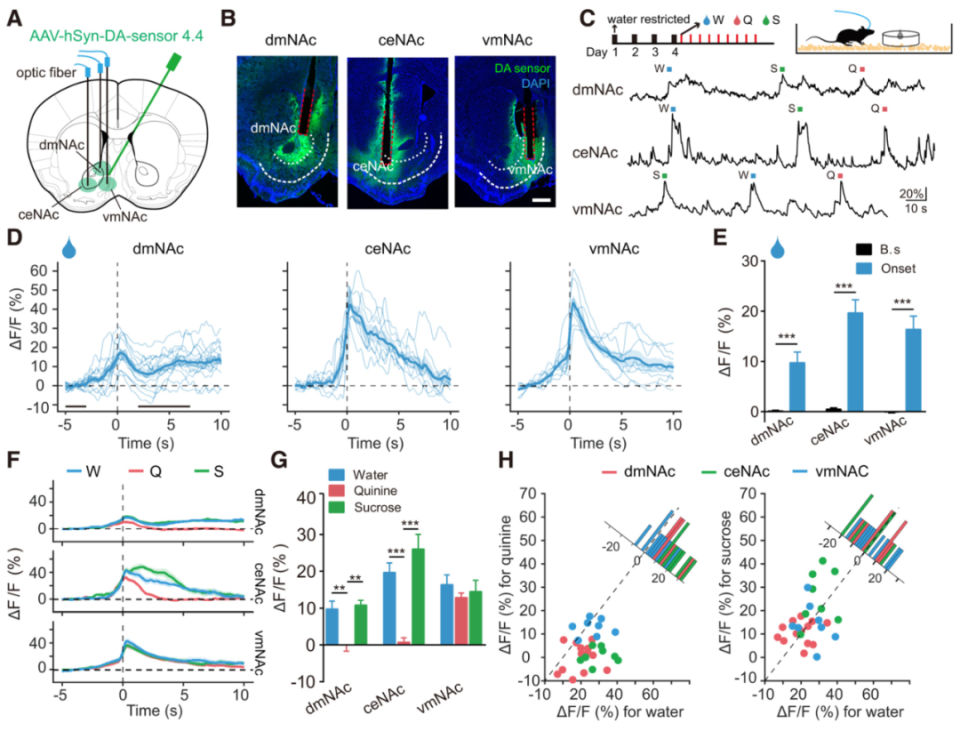
说起风情,少不了多巴胺相关奖赏系统。多巴胺(DA)传递是奖赏过程的核心,包括动机和强化。动机是诱发某种行为的内在过程,而强化通过刺激–反应和行动–结果的关联增加行为发生概率。这两种功能被协调起来以最大化奖赏结果,与多种神经精神疾病(如物质使用障碍和情绪障碍)密切相关。
腹侧纹状体壳区(mNAcSh)是奖赏系统关键核团,接受来自腹侧被盖区(VTA)的多巴胺能输入。多项研究指出,多巴胺的释放过程很可能是动机和强化的基础。然而,多巴胺释放如何通过DA受体行使其功能,进而驱动奖赏功能的不同方面,尚未可知。
mNAcSh内中型多棘神经元(MSNs)高表达多巴胺受体,包括D1受体(D1R)和D2受体(D2R),D1-MSN与D2-MSN基本不共标。多巴胺会作用于D1R和D2R,对D1-MSNs和D2-MSNs产生相反效应。此外,mNAcSh还富含D3受体(D3R),是一种Gαi/o偶联的G蛋白偶联受体,属于D2类受体家族。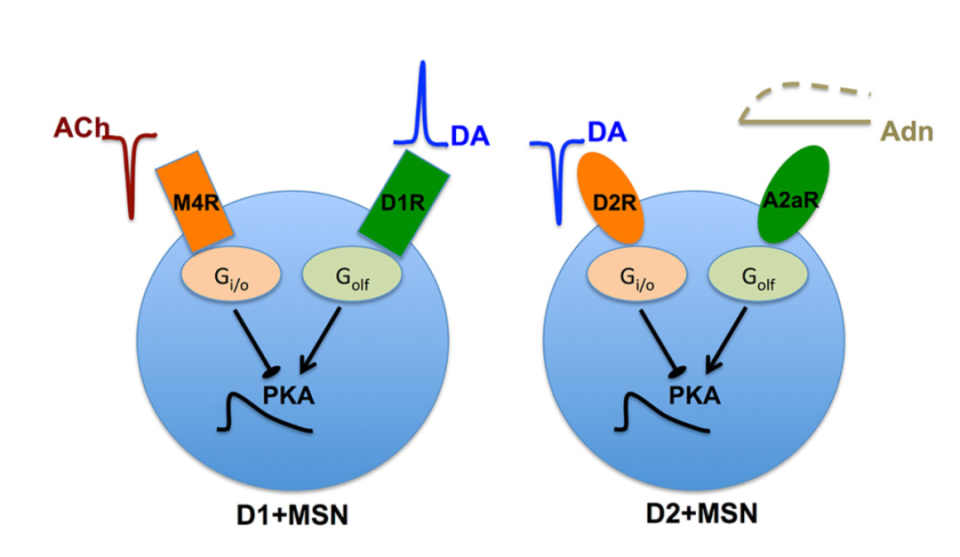
图片来源: Nair et al.,J Neurosci.
D3R是一种高亲和力多巴胺受体,其亲和力比D2R高十倍。药理学阻断伏隔核(NAc)D3R信号传导可抑制药物渴求行为。然而, D3R拮抗剂亦可能对D2R产生非特异性作用。另有研究发现,全身过表达或敲除D3R影响机体对可卡因的易感性与动机,但这些研究并未特异性针对mNAcSh D3R,而非伏隔核D3R信号传导也可能参与奖赏调节。因此,缺乏选择性操控mNAcSh D3R表达水平的工具限制了科研工作者对其在奖赏调节中的贡献的理解。有趣的是,伏隔核核心/壳区的D1-MSNs共表达D3R与D1R。由于D3R和D1R对多巴胺的亲和力不同,且诱发功能相反的信号效应器(分别为Gαi/o和Gαs/olf蛋白),让mNAcSh内多巴胺信号的作用机制更加错综复杂。
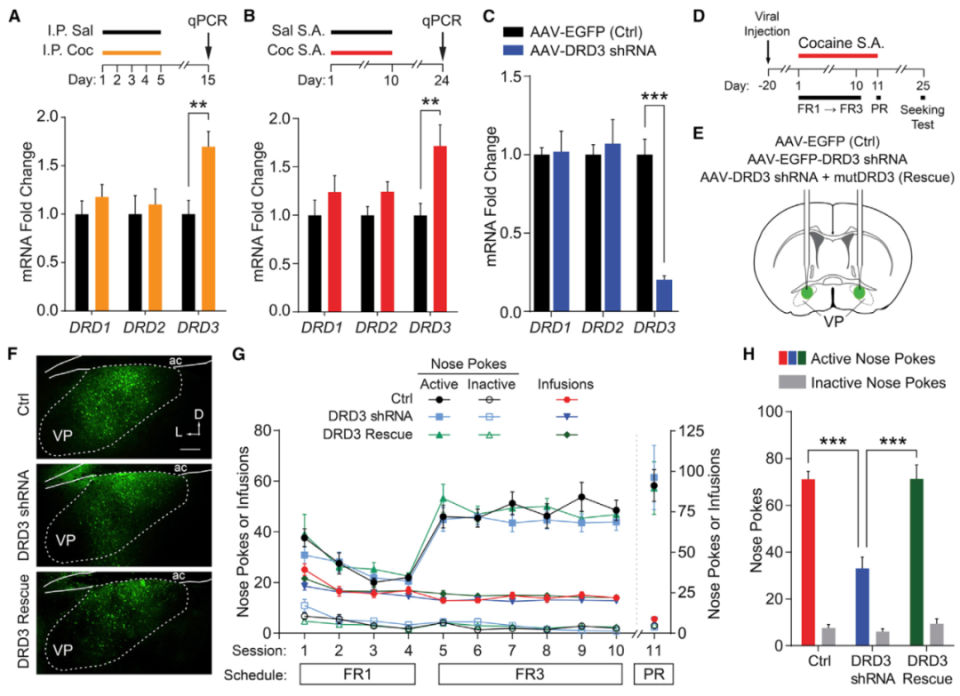
2024年12月9日,《Nature Neuroscience》杂志在线刊登了美国NIH Hugo A. Tejeda 课题组的最新重要工作,他们结合神经示踪、膜片钳电生理以及神经环路水平的DA受体操控手段,发现mNAcSh D3R的缺失会影响动机,但不影响条件反射或负性强化过程。相反,敲除D1R会影响条件反射和负性强化,却不影响动机。此外,作者还证实D3R和D1R在调节MSN突触功能方面扮演不同角色。此研究揭示了mNAcSh内D3R和D1R在奖赏功能调控中的不同作用,进一步深化了科研工作者对动机与强化的理解,对探索神经精神疾病的潜在治疗靶点具有重要意义。

首先,为探究mNAcSh内D3R功能,作者在Drd3fl/fl小鼠mNAcSh内双侧注射AAV-GFP-Cre。敲除mNAcSh内D3R后,小鼠在wheel running、running disk choice、lever press (for chocolate)范式的动机水平均显著下调(图1),而OFT内运动水平、焦虑水平及糖水偏好均不受影响(补充材料1),提示mNAcSh内D3R对动机是必要的,而对运动、情绪及liking行为不是。
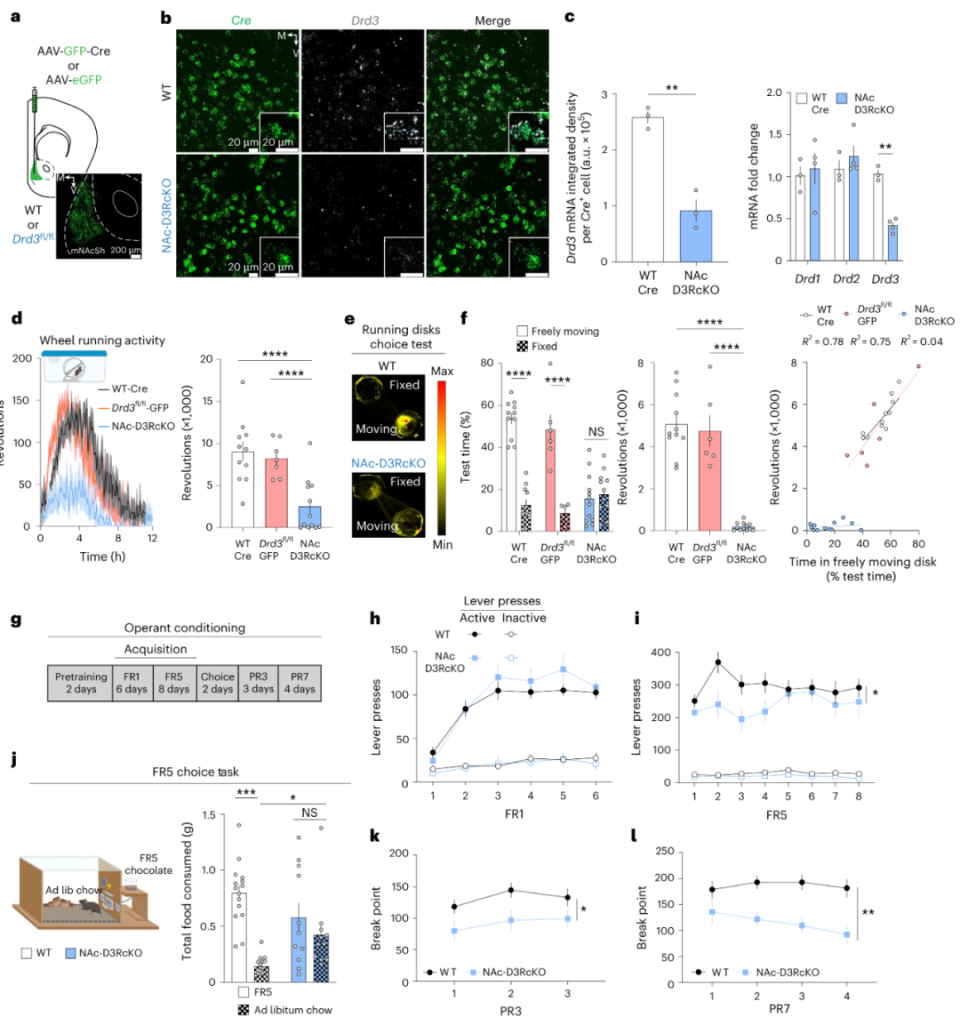
图1 敲除mNAcSh内D3R引起动机缺失
接下来,回答以上问题,作者行原位杂交实验,发现D3R主要与D1R共标,与D2R也有少量共标(图2a-f)。研究表明,D1-MSNs投射到VP、LH和VTA,而D2-MSNs特异性投射到VP。通过神经顺向示踪实验,作者发现D3R阳性神经元投射到VP、LH和VTA。为进一步研究mNAcSh D3R阳性神经元的功能性连接,作者结合光遗传学与膜片钳电生理,发现mNAcSh D3R阳性神经元与VP、LH和VTA神经元形成抑制性突触联系,其中到VP的投射最强(图2g-k)。再然后,作者结合逆向示踪与原位杂交实验。有趣的是,投射到VP的mNAcSh D3R阳性神经元比率最低(图2l-n)。以上,便是mNAcSh D3R阳性神经元的解剖学性质。那么,哪条通路参与动机行为呢?
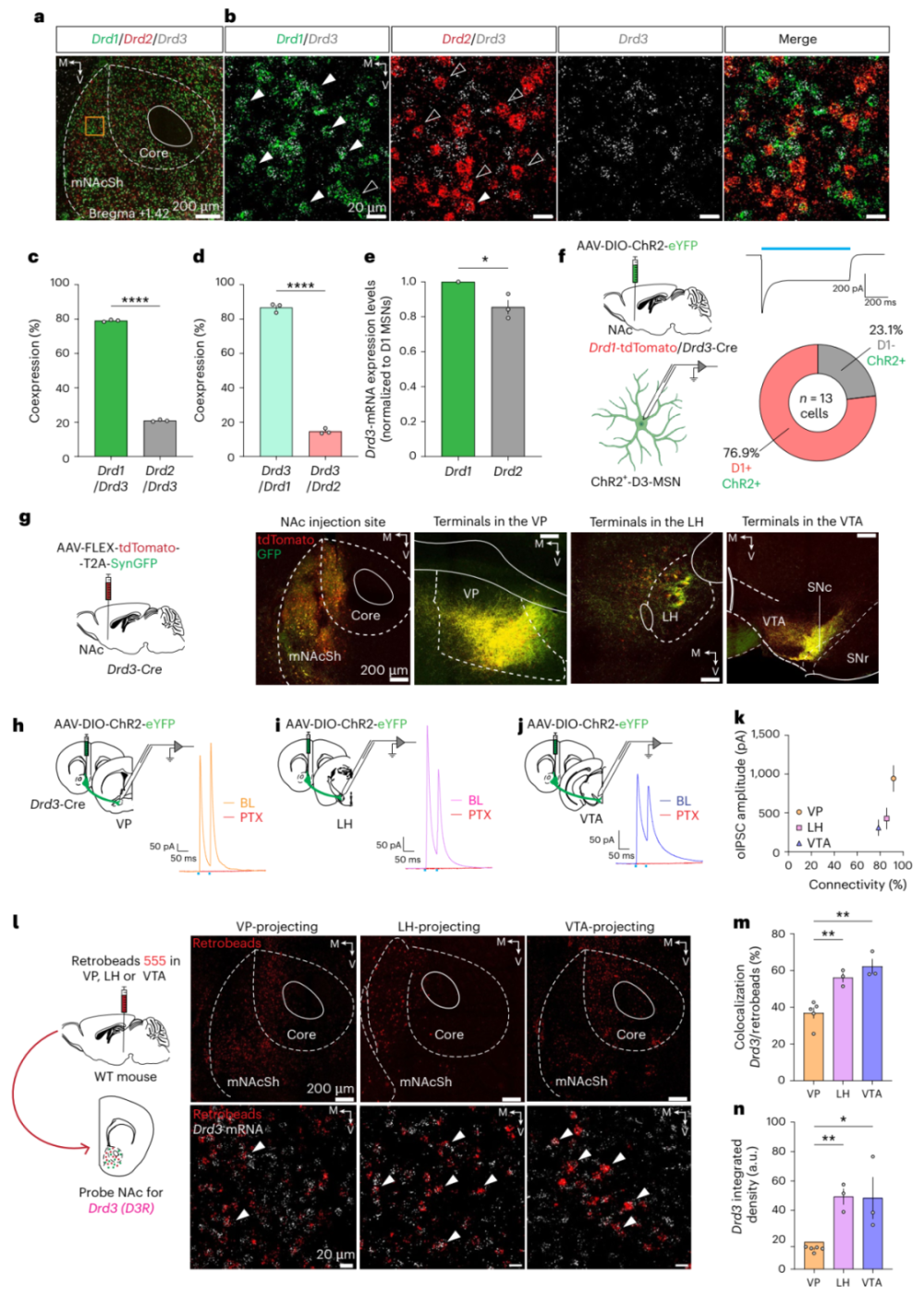
图2 mNAcSh内D3R主要表达于D1-MSNs
为回答以上问题,作者结合逆向标记、双重组酶与基因特异性敲除手段,在Drd3fl/fl小鼠的VP、LH或VTA内注射CAV-Flp,在NAcSh内注射AAV-fDIO-Cre。令人惊讶的是,敲低投射至VP、LH或VTA的mNAcSh神经元内D3R均显著降低小鼠在wheel running、running disk choice范式的的动机行为(图3a-h)。对PR7 lever press范式,敲低投射至VTA的mNAcSh神经元内D3R不影响动机行为,其他两种略有影响(图3i-k)。以上结果表明,投射至不同下游的mNAcSh神经元内D3R均影响动机行为。我们知道,多种脑功能与突触可塑性密切相关。那么,D3R对mNAcSh神经元不同下游的突触可塑性影响如何呢?
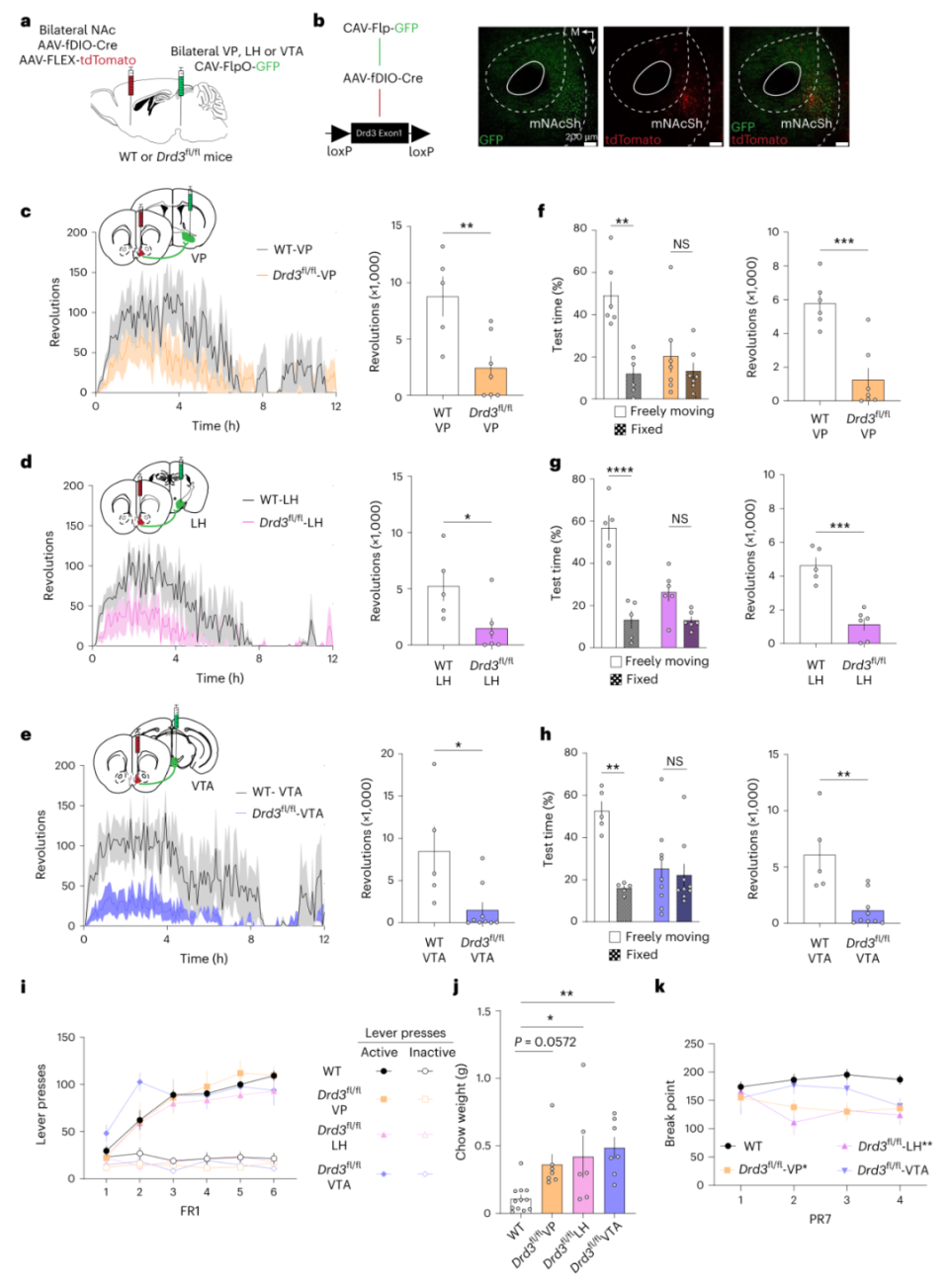
图3 敲除mNAcSh内D3R引起动机缺失无下游特异性
突触前D3R选择性抑制mNAcSh D3R-VP通路的GABA能突触联系
研究发现,Gi/o偶联GPCR(包括D3R)通过抑制轴突末梢突触递质释放以影响突触可塑性。为进一步探究mNAcSh D3R的功能,作者再次采用膜片钳电生理手段,并引入Cre-ON(DIO-ChR2)与Cre-OFF(DO-ChR2)系统。他们在Drd1-tdTomato/Drd3-Cre小鼠mNAcSh内注射AAV-DIO/DO-ChR2,记录D1-MSN(tdTomato+)或D2-MSN(tdTomato-)。D3R激动剂PD-128901减弱D3R阳性神经元,而非D3R阴性神经元,到D1-MSN与D2-MSN的抑制性突触连接(图4a-e;补充材料6)。然后,是上文三个下游。其中,只有mNAcSh D3R阳性神经元到VP的抑制性突触联系受PD-128901削弱,LH与VTA神经元oIPSC幅度与PPR不受影响(图4f-q),提示mNAcSh D3R-LH/VTA突触前膜缺乏D3R表达。图4 突触前D3R选择性抑制mNAcSh D3R-VP通路的GABA能突触联系上文已知,投射到VP、LH和VTA的mNAcSh神经元内D3R均参与动机行为,但只有mNAcSh-VP的突触前有D3R表达。由此,作者推测mNAcSh内,而非投射到其他脑区的轴突末梢,的D3R对动机行为至关重要。为验证此假设,作者首先需要D3R拮抗剂SB-277011A,并探究其功能。他们发现双侧mNAcSh给予SB-277011A的确可以降低小鼠动机行为(图5a-c)。他们在Drd3fl/fl小鼠mNAcSh单侧注射AAV-Cre以敲除单侧D3R,对照组在同侧给予D3R拮抗剂SB-277011A,实验组在对侧给予SB-277011A。如此,对照组一侧mNAcSh的D3R功能受损,另一侧mNAcSh的D3R功能保持完整,此状态下小鼠同样具有动机行为;而实验组一侧mNAcSh的D3R功能受损,另一侧mNAcSh内部D3R功能受损,但所有下游突触前的D3R功能完好,此状态下小鼠动机水平显著降低(图5d-g),表明mNAcSh内部D3R才是动机行为的核心,突触前的D3R可能没那么重要。笔者注:置管给予药物和AAV基因敲除最大的区别就是,置管只针对某个脑区,而AAV基因敲除针对某脑区及所有下游的突触前;当然,并不能说置管更优,因为置管给予药物还会影响其他脑区投过来的突触前,而AAV基因敲除则不会。
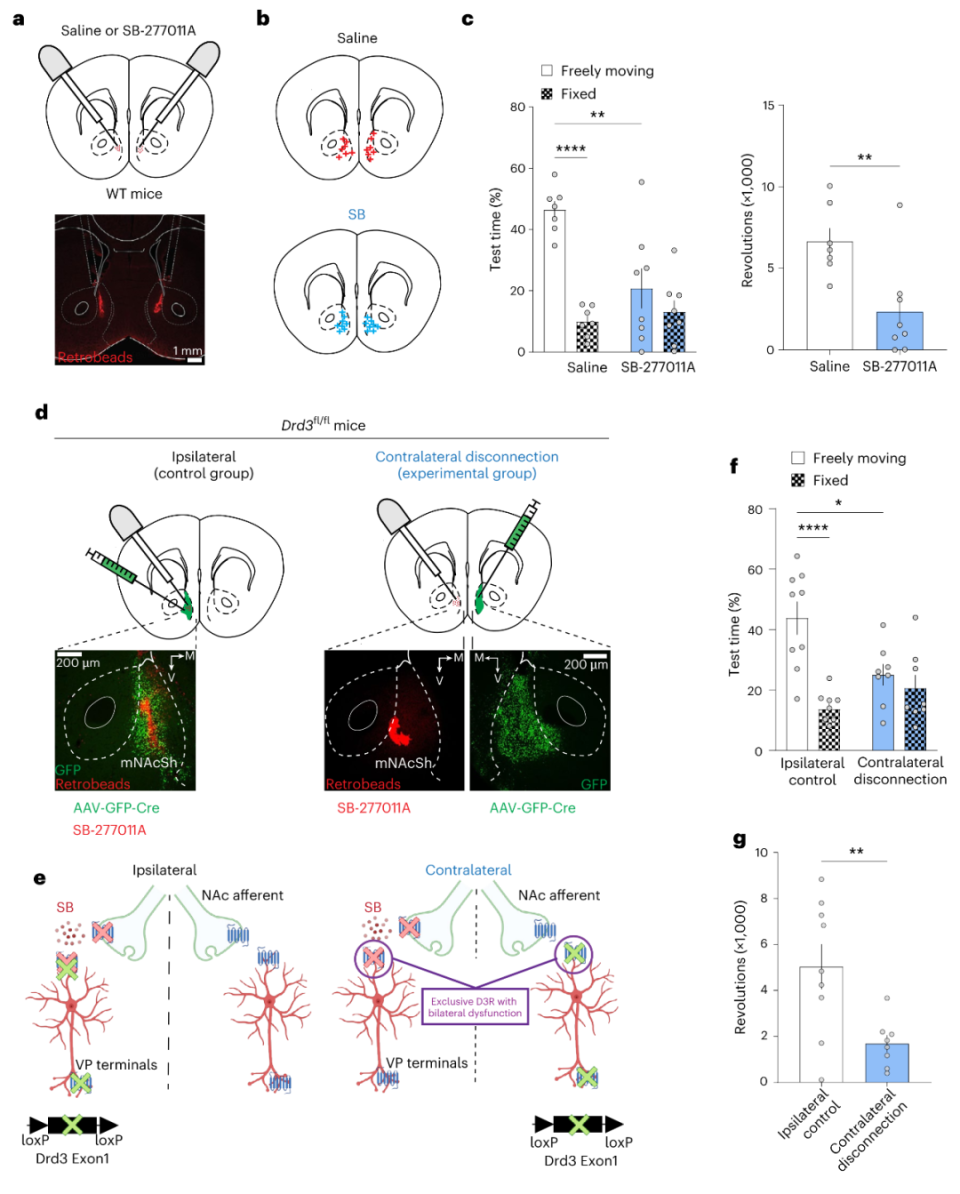
图5 mNAcSh内部的D3R信号对动机行为至关重要为探究D1R功能,作者引入Drd1fl/fl小鼠。双侧敲除mNAcSh D1R显著降低Lever press(FR1)范式内小鼠的压杆次数,但其他动机行为不受影响,糖水偏好、社交、黑白箱、新奇物体识别亦不受影响;而敲除mNAcSh D3R不影响Lever press(FR1)范式内小鼠的压杆次数(图6a-g),提示mNAcSh D1R参与的是action-outcome contingencies相关的强化过程,而不参与动机行为。为进一步验证此结论,作者采用基于CS-US的Platform-mediated avoidance范式。双侧敲除mNAcSh D1R显著降低小鼠的逃跑行为,但对奖赏性CS-US的强化过程不受影响,敲除mNAcSh D3R均无明显差异(图6h-l)。综上,mNAcSh D3R介导动机而非强化,mNAcSh D1R介导强化而非动机,且并不参与所有类型动机。既然二者均表达于D1-MSNs,那么D3R与D1R的功能如何协调呢?
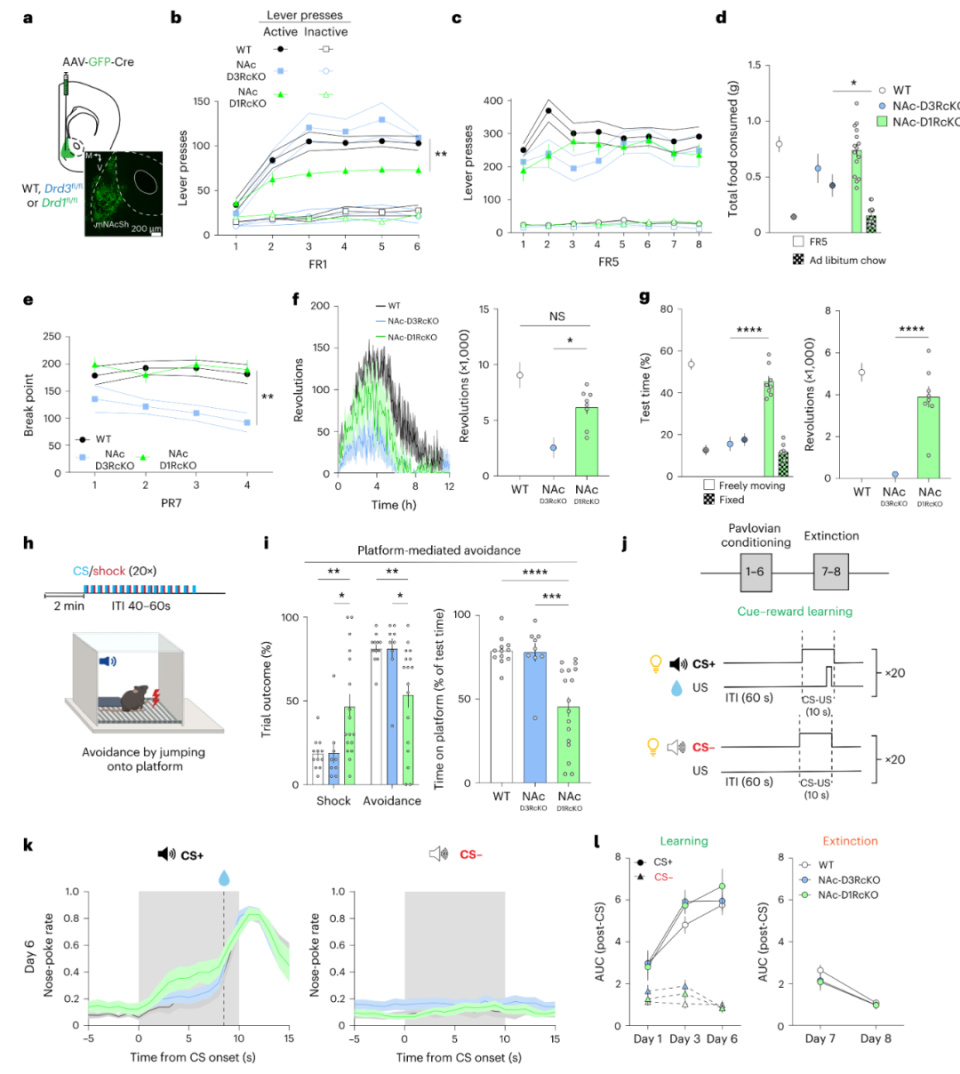
研究表明,奖赏性学习(强化)与NMDAR相关的突触可塑性关系重大,于是作者行膜片钳电生理记录D1-MSNs,通过紫外诱导MNI-glutamate释放以诱发NMDAR电流。施加D1R激动剂SKF-81297可增加NMDAR电流幅度,施加D1R拮抗剂SCH-31966可阻断此过程,施加D3R激动剂则无此现象(图7a-d),敲除D1R后亦无此现象(图7e-h)。对于mNAcSh D3R阳性神经元与D1-MSNs的GABA能突触联系,D1R激动剂并不影响其突触强度,D3R激动剂降低其突触强度(图7i-l)。最后,作者探究mNAcSh内部D1R是否对强化行为是必要的。他们再次采用图5的方法,得到了肯定答案(图7m-n)。综上所述,生理学水平上,D1R与D3R在D1-MSNs内扮演不同角色,进而介导奖赏功能的不同成分——动机与强化。
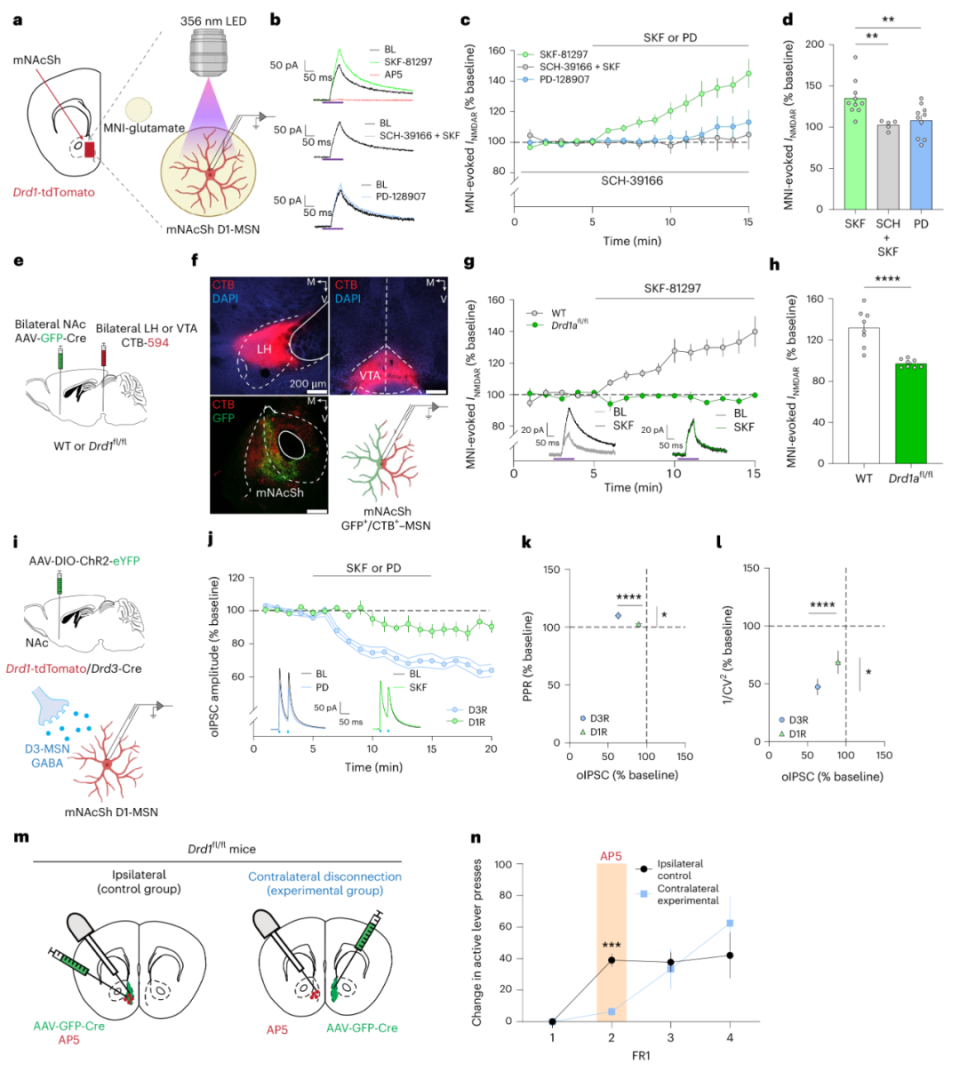
图7 mNAcSh内部的D1R信号对强化行为至关重要
本篇文章十分精彩,但并非完美无瑕:1、方法方面,作者注射病毒的速度为100nl/min,这种高速注射可能会造成神经元死亡,进而影响行为学结果。2、机制方面,作者选择了VP、LH、VTA作为下游,表示mNAcSh D3R神经元不具有下游特异性;但mNAcSh还有其他很多下游,也许投射到其他下游的mNAcSh D3R神经元功能会有差异。3、作者通过单侧基因敲除+单侧药理学证实mNAcSh D3R的重要性,不代表mNAcSh-VP突触前的D3R不参与动机行为。4、此外,不同下游如何参与动机与强化,值得进一步探究。mNAcSh内多巴胺释放能够表征奖励的不同特征,例如动机和强化。然而,多巴胺受体如何将DA的释放转化为奖励的不同构成要素,其分子、细胞、环路机制尚不清楚。在本研究中,作者发现mNAcSh中D3R的信号通过调节局部微回路,促进小鼠动机行为。D3R与D1R共表达于D1-MSNs,而D1R主要调节强化过程,并不参与动机行为。此外,D3R和D1R信号在mNAcSh神经元中的生理学功能亦不相同。本篇文章的研究结果建立了一个基本框架,即在同一类mNAcSh神经元中,多巴胺信号通过不同多巴胺受体的作用在生理功能上实现特异性。这种结构和功能的组织方式,使边缘系统神经元能够协调奖赏相关行为的不同方面,可能与动机与强化相关神经疾病诱因密切相关。Enriquez-Traba, J., Arenivar, M., Yarur-Castillo, H.E. et al. Dissociable control of motivation and reinforcement by distinct ventral striatal dopamine receptors. Nat Neurosci (2024). https://doi.org/10.1038/s41593-024-01819-9