肝癌是全球死亡率极高、年发病率呈上升趋势且预后差的肿瘤,采用经典疗法治疗后易出现术后并发症、化疗药耐药等情况。肿瘤免疫疗法可通过增强机体固有免疫和/或适应性免疫,从而提高其对肿瘤细胞的控制和杀伤能力,其中树突状细胞(dendritic cell,DC)疫苗在肿瘤治疗中发挥了重要作用。DC是一类多功能且最有效的抗原提呈细胞(antigen presenting cell,APC),具有强大的免疫活性,根据其活化成熟程度分为未成熟DC(immature DC,imDC)和成熟DC(mature DC,mDC);imDC的抗原摄取加工能力虽强,但抗原提呈及激活细胞毒性T淋巴细胞(简称“T细胞”)能力有限,而mDC不仅抗原提呈能力强,还能提供T细胞激活所需的所有信号。已有研究发现,处于肿瘤环境中的mDC数量不足及其抗原提呈功能缺陷、imDC过多聚集会导致DC疫苗达不到预期疗效。信号转导和转录激活因子4(signal transducer and activator of transcription-4,STAT-4)能诱导imDC成熟并增强其对T细胞的激活能力,而激活Toll样受体/核转录因子κB(nuclear factor-kappa B,NF-κB)信号通路则能够上调STAT-4表达。因此,研发促进imDC成熟、改善并提高mDC抗原提呈功能的药物,对于提高DC疫苗作用具有重要意义。
多糖是新疆特色药用植物胀果甘草Glycyrrhiza inflata Batal.中具有免疫调节作用的活性成分。本课题组前期研究发现,胀果甘草粗多糖(G. inflata polysaccharides,GiP)及其纯化多糖GiP-B1可在体外通过激活Toll样受体/NF-κB信号通路来促进imDC成熟,提高白细胞介素12(interleukin-12,IL-12)分泌和抗原提呈功能,从而发挥增强免疫功能的作用。研究发现,GiP及GiP-B1佐助的DC疫苗对肝癌细胞H22荷瘤小鼠具有抗肿瘤作用,但其作用机制是否与GiP、GiP-B1调节肿瘤中DC的功能有关尚不明确。为此,本研究探讨GiP及GiP-B1对荷瘤小鼠imDC促成熟的影响,并通过共培养体系评价GiP及GiP-B1促成熟的荷瘤小鼠mDC以T细胞为介导的抗肿瘤能力以及作用机制,以期为开发胀果甘草多糖-DC疫苗提供理论基础和实验依据。HERACELL 150i型CO2恒温细胞培养箱、Multiskan GO型全波长酶标仪、NanoDrop2000/200C型核酸蛋白定量仪均购自美国Thermo Fisher Scientific公司;AB135-S型电子分析天平购自瑞士Mettler Toledo公司;3-18K型低温离心机购自德国Sigma公司;HH-S4型恒温水浴锅购自江苏金怡仪器科技有限公司;LSRFortessa型流式细胞仪购自美国BD公司;0.4 μm孔径Transwell嵌套装置购自北京兰杰柯科技有限公司;QuantStudio 6型实时定量聚合酶链式反应(polymerase chain reaction,PCR)系统购自美国ABI公司;DYCZ-24DN型电泳仪购自北京六一生物科技有限公司;SH-523型化学发光成像系统购自杭州申花科技有限公司。
GiP由本课题组前期按照文献方法从胀果甘草G. inflata Bat.中提取所得,含量为52.66%;通过柱层析法进行纯化、精制得到GiP-B1,含量为100%,分子量约为2.0×106 Da,具有聚鼠李糖-半乳糖醛酸主链。重组小鼠肿瘤坏死因子α(tumor necrosis factor-α,TNF-α)(批号061454-1)购自美国PeproTech公司;小鼠骨髓imDC完全培养基(批号CM-M151A)购自武汉普诺赛生命科技有限公司;RPMI-1640培养基、青霉素-链霉素混合液(双抗)、磷酸盐缓冲液(phosphate buffer saline,PBS)(批号分别为2152424、2245296、2201008)均购自上海龙田生物科技有限公司;胎牛血清(批号42G5093K)购自美国Gibco公司;CCK-8试剂盒(批号PM508)购自日本同仁化学研究所;抗小鼠的mDC表面标志物CD11c、CD86、CD80、MHC-Ⅱ单克隆抗体(批号分别为B383301、B318893、B321317、B336920)均购自美国BioLegend公司;CD4+T细胞磁珠分选试剂盒(批号130-104-454)购自德国Miltenyi Biotec公司;BCA蛋白定量试剂盒、小鼠脾淋巴细胞分离试剂盒、RIPA组织/细胞裂解液(批号分别为PC0020、P8860、R0010)均购自北京索莱宝科技有限公司;乳酸脱氢酶(lactate dehydrogenase,LDH)细胞毒性检测试剂盒(批号C0017)购自上海碧云天生物技术股份有限公司;小鼠IL-12p70、干扰素-γ(interferon-γ,IFN-γ)、IL-4、IL-10酶联免疫吸附测定实验(ELISA)试剂盒(批号分别为EK0422、EK0375、EK0405、EK0417)和辣根过氧化物酶标记的羊抗兔IgG二抗(批号BA1054)均购自武汉博士德生物工程有限公司;细胞总RNA提取试剂盒(批号C0017)购自成都福际生物技术有限公司;SYBR Green Mix荧光定量试剂、反转录试剂盒(批号分别为Q111-02、R223-01)均购自南京诺唯赞生物科技股份有限公司;兔抗小鼠NF-κB p65、STAT-4单克隆抗体(批号分别为ab32536、ab284408)均购自英国Abcam公司;兔抗小鼠IL-12受体β2(IL-12Rβ2)单克隆抗体(批号A10348)购自武汉爱博泰克生物科技有限公司;兔抗小鼠磷酸化NF-κB p65(phosphorylated NF-κB p65,p-NF-κB p65)、磷酸化STAT-4(phosphorylated STAT-4,p-STAT-4)多克隆抗体(批号分别为AF2006、AF3441)均购自江苏亲科生物研究中心有限公司;小鼠抗β-肌动蛋白(β-actin)单克隆抗体(批号66009-1-Ig)购自武汉三鹰生物技术有限公司。本研究所用动物为健康C57BL/6雄性小鼠与昆明种雌性小鼠,SPF级,6~8周龄,体重18~20 g,分别购自新疆医科大学动物实验中心、湖南斯莱克景达实验动物有限公司、北京维通利华实验动物技术有限公司,实验动物生产许可证号分别为SCXK(新)2018-0002、SCXK(湘)2019-0004、SCXK(京)2021-0006。本研究得到新疆医科大学实验动物伦理委员会审批,伦理审批号为IACUC-20210405-1。1.4 细胞株 小鼠肝癌细胞H22(批号CL-0341)购自武汉普诺赛生命科技有限公司。 2.1 H22细胞培养上清液及H22冻融抗原的制备 用含10%胎牛血清、1%双抗的RPMI-1640完全培养基,在37 ℃、5%CO2培养箱中培养H22细胞,待细胞长至80%~90%时即可传代。待H22细胞呈对数生长期时,收集细胞,离心,取上清液置于-80 ℃冰箱中保存,备用。取细胞沉淀,用RPMI-1640完全培养基重悬,调整密度为1×107个/mL,反复冻融5次,离心,取上清液即为H22冻融抗原;留少许冻融抗原置于冰上测抗原浓度,其余冻融抗原置于-80 ℃冰箱中保存,备用。取对数生长期的H22细胞,离心,取细胞沉淀,用PBS调整密度为1×107个/mL,然后取5只健康昆明种雌性小鼠,均于腹腔接种0.2 mL以保种传代。腹腔传代第5天,抽取乳白色腹水,洗涤,重悬,再次接种到另外5只健康昆明种雌性小鼠腹腔,连续传3代,抽取腹腔生长旺盛的腹水H22细胞,洗涤,用PBS调整密度为1×107个/mL。随后取12只健康C57BL/6雄性小鼠,均于右腋皮下注射该细胞悬液0.1 mL,以腋下形成黄豆粒大小的包块作为荷瘤小鼠造模成功。将造模成功的荷瘤小鼠颈椎脱臼处死,在无菌条件下取出股骨和胫骨,剪去骨头两端,用RPMI-1640基础培养基冲洗骨髓腔内细胞至骨髓变白,然后反复吹打骨髓细胞悬液,过滤,洗涤,离心,取上清液用imDC完全培养基重悬细胞,在CO2恒温培养箱中静置培养48 h后,用含20%H22细胞培养上清液的imDC完全培养基全量换液,第4天半量换液,培养至第6天的细胞即为荷瘤小鼠imDC。2.3 GiP及GiP-B1对荷瘤小鼠imDC促成熟作用的研究 2.3.1 GiP及GiP-B1给药浓度和作用时间的筛选 采用CCK-8法进行检测。取“2.2”项下荷瘤小鼠imDC,以2×104个/孔接种于96孔板中,分为对照组、TNF-α组(20 ng/mL)、GiP及GiP-B1不同质量浓度组(50、100、200、400 μg/mL),每组设3个复孔。各孔加入含或不含相应药物的imDC完全培养基培养24、48 h,培养结束前4 h,各孔加入4 μg/mL H22冻融抗原;培养结束后,加入含10%CCK-8试剂的培养基,于37 ℃避光孵育2 h。采用酶标仪测定450 nm波长处各孔mDC的光密度(optical density,OD)值,计算细胞活力:细胞活力(%)=(实验组OD值-培养基对照的OD值)/(对照组OD值-培养基对照的OD值)×100%。根据检测结果筛选GiP及GiP-B1的最佳给药浓度和作用时间。2.3.2 荷瘤小鼠mDC中表面标志物阳性表达率的检测 采用流式细胞术进行检测。取“2.2”项下荷瘤小鼠imDC,以1×106个/孔接种于6孔板中,按照“2.3.1”项下方法分组、给药(GiP及GiP-B1的给药浓度均100 μg/mL),作用时间为24 h(根据“2.3.1”项下结果设置)。培养结束后,收集细胞,离心,弃上清液,用PBS洗涤2次,重悬,分别加入CD11c、CD80、CD86、MHC-Ⅱ单克隆抗体各2 μL,4 ℃下避光孵育30 min;用PBS洗去未结合的抗体,离心,弃上清液;细胞用PBS重悬,采用流式细胞仪和Flow Jo 10.8.1软件检测细胞中CD11c、CD80、CD86、MHC-Ⅱ阳性表达率。2.3.3 荷瘤小鼠mDC上清液中IL-12p70、IL-4水平的检测 采用ELISA法进行检测。按照“2.3.2”项下方法分组、给药、培养后,收集细胞培养上清液,按照ELISA试剂盒说明书方法操作,采用酶标仪检测上清液中 IL-12p70、IL-4水平。2.4 GiP及GiP-B1促成熟的荷瘤小鼠mDC的抗肿瘤作用及机制研究2.4.1 荷瘤小鼠mDC刺激CD4+T细胞增殖的作用及二者共培养比例的筛选 在无菌条件下取出健康C57BL/6雄性小鼠脾脏,分离脾淋巴细胞,借助磁珠分选试剂盒筛选、纯化后得CD4+T细胞。取荷瘤小鼠imDC,按照“2.3.2”项下方法分组、给药、培养后,离心,收集细胞(即荷瘤小鼠mDC)。取荷瘤小鼠mDC与CD4+T细胞,以每孔各100 μL接种于96孔板中,二者比例分别为1∶5、1∶10、1∶20、1∶40(以细胞数量计,下同),每组设3个复孔;同时设置刺激细胞对照组(只接种荷瘤小鼠mDC,用于观察细胞活力)与反应细胞对照组(只接种CD4+T细胞)。共培养24 h后生成CD4-细胞毒性T细胞(CD4-CTL),加入含10%CCK-8试剂的培养基,于37 ℃避光孵育2 h。采用酶标仪检测450 nm波长处各孔的OD值,计算刺激指数:刺激指数=(实验组OD值-培养基对照的OD值)/(反应细胞对照组OD值-培养基对照的OD值)。刺激指数越大,CD4+T细胞增殖越明显,说明经GiP及GiP-B1促成熟的mDC具有刺激活化T细胞的能力。根据检测结果筛选荷瘤小鼠mDC与CD4+T细胞共培养的最佳比例。2.4.2 CD4-CTL上清液中IL-12p70、IFN-γ、IL-4、IL-10水平的检测采用ELISA法进行检测。按照“2.4.1”项下方法分组、给药、共培养,荷瘤小鼠mDC与CD4+T细胞共培养的比例为1∶5(根据“2.4.1”项下结果设置)。共培养24 h后,收集细胞培养上清液,按照ELISA试剂盒说明书方法操作,采用酶标仪检测上清液中IL-12p70、IFN-γ、IL-4、IL-10水平。2.4.3 CD4-CTL对H22细胞的杀伤活性检测按照“2.4.2”项下方法分组、给药、共培养24 h后,收集悬浮细胞(即CD4-CTL)。取CD4-CTL(效应细胞)和H22细胞(靶细胞),以每孔各100 μL接种于96孔板中,效应细胞与靶细胞比(以下简称“效靶比”)分别为10∶1、20∶1、50∶1(以细胞数量计),培养24 h,每组设3个复孔;同时设置各效靶比对应的效应细胞自然释放组(只加效应细胞)、靶细胞自然释放组(只加靶细胞)、靶细胞最大释放组(加靶细胞和LDH释放剂)。培养结束前1 h,在靶细胞最大释放组中加入LDH释放剂2 μL,于37 ℃避光孵育1 h。培养结束后,离心,吸出各孔等体积上清液,加入到新的96孔板中,每孔加入LDH工作液60 μL,混匀,室温下避光孵育30 min;采用酶标仪检测490 nm波长处各孔的OD值,计算杀伤活性:杀伤活性(%)=(实验组OD值-效应细胞自然释放组OD值-靶细胞自然释放组OD)/(靶细胞最大释放组OD值-靶细胞自然释放组OD)×100%。2.4.4 共培养后荷瘤小鼠mDC中IL-12、IL-12R、STAT-4 mRNA表达检测 采用实时定量PCR法进行检测。取荷瘤小鼠imDC,按照“2.3.2”项下方法分组、给药、培养后,离心,取荷瘤小鼠mDC接种于Transwell嵌套装置上室,将“2.4.1”项下纯化后得到的CD4+T细胞接种于下室,共培养比例为1∶5(根据“2.4.1”项下结果设置)。共培养24 h后,收集并提取Transwell装置上室内各组mDC总RNA,反转录为cDNA,以上述cDNA为模板进行PCR扩增。反应体系(20 μL)包含cDNA模板4 μL,SYBR Green Master Mix 10 μL,正、反向引物各0.4 μL,50×ROX Reference Dye 2 0.4 μL,ddH2O 4.8 μL。反应条件为:95 ℃预变性10 min;95 ℃变性15 s,60 ℃退火延伸60 s,共40个循环。以甘油醛-3-磷酸脱氢酶(GAPDH)为内参,采用2-ΔΔCt法计算IL-12、IL-12R、STAT-4 mRNA表达量。引物由擎科生物科技(北京)股份有限公司设计、合成,其序列及产物长度见表1。
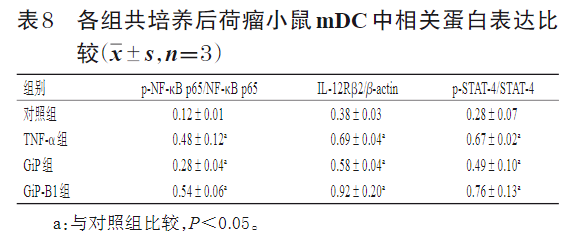
2.4.5 共培养后荷瘤小鼠mDC中IL-12/NF-κB/STAT-4信号通路相关蛋白表达检测 采用Western blot法进行检测。取荷瘤小鼠mDC按照“2.4.4”项下方法分组、给药、共培养24 h后,取Transwell装置上室内各组mDC,充分裂解,离心,取上清液,测定蛋白浓度后煮沸变性。取变性蛋白样品适量,经电泳分离后转膜,封闭,加入β-actin、NF-κB p65、p-NF-κB p65、IL-12Rβ2、STAT-4、p-STAT-4一抗(稀释比例分别为 1∶5 000、1∶50 000、1∶1 000、1∶1 000、1∶1 000、1∶1 000),于4 ℃孵育过夜;加入相应二抗(稀释比例为1∶10 000),室温下孵育2 h;以化学发光法显色,采用Image-Pro Plus 6.0软件分析蛋白条带。以目的蛋白与内参蛋白(β-actin)的灰度值比值表示目的蛋白的表达量,再进一步以p-NF-κB p65与NF-κB p65、p-STAT-4与STAT-4的蛋白表达量比值表示NF-κB、STAT-4蛋白的磷酸化水平。
采用SPSS 25软件对数据进行统计分析。数据以x±s表示,多组间比较采用单因素方差分析,组间两两比较采用LSD-t检验。检验水准α=0.05。
3.1 GiP及GiP-B1对荷瘤小鼠imDC的促成熟作用 3.1.1 GiP及GiP-B1的给药浓度和作用时间 经100 μg/mL GiP,50、100 μg/mL GiP-B1干预24 h后,mDC细胞活力分别为(133.45±15.16)%、(126.50±13.17)%、(132.49±12.05)%(n=3),均显著高于对照组(细胞活力为100%)(P<0.05),其中100 μg/mL GiP-B1组的mDC细胞活力显著高于50 μg/mL GiP-B1组(P<0.05)。其余各组mDC细胞活力与对照组比较,差异均无统计学意义(P>0.05)。故选择后续实验GiP及GiP-B1的给药浓度为100 μg/mL,作用时间为24 h。3.1.2 GiP及GiP-B1对荷瘤小鼠mDC中表面标志物阳性表达率的影响 与对照组比较,TNF-α组、GiP组与GiP-B1组荷瘤小鼠mDC中MHC-Ⅱ阳性表达率均显著升高(P<0.05),CD11c、CD80、CD86阳性表达率均有升高趋势,但与对照组比较差异均无统计学意义(P>0.05)。结果见表2、图1。
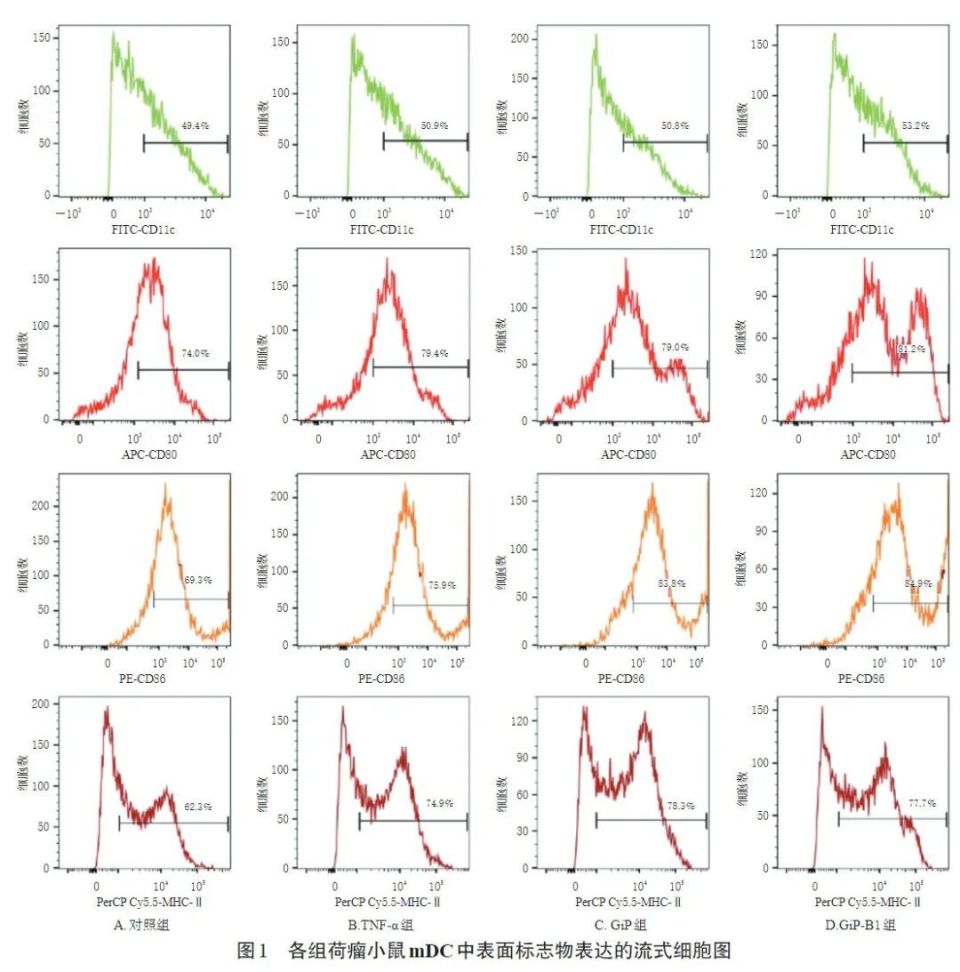
3.1.3 GiP及GiP-B1对荷瘤小鼠mDC上清液中细胞因子水平的影响 与对照组比较,GiP组与GiP-B1组荷瘤小鼠mDC上清液中IL-12p70、IL-4水平以及TNF-α组小鼠mDC上清液中IL-4水平均显著升高(P<0.05)。结果见表3。
3.2 GiP及GiP-B1促成熟的荷瘤小鼠mDC的抗肿瘤作用及作用机制 3.2.1 荷瘤小鼠mDC刺激CD4+T细胞增殖的作用及二者共培养的最佳比例 荷瘤小鼠mDC与CD4+T细胞的比例分别为1∶5、 1∶10、1∶20、1∶40时,与对照组比较,TNF-α组、GiP组及GiP-B1组荷瘤小鼠mDC刺激CD4+T细胞增殖的刺激指数均显著升高(P<0.05),其中荷瘤小鼠mDC与CD4+T细胞的比例为1∶5时各组的刺激指数最大且CD4+T细胞增殖最显著(表4)。故选择后续实验荷瘤小鼠mDC与CD4+T细胞的共培养比例为1∶5。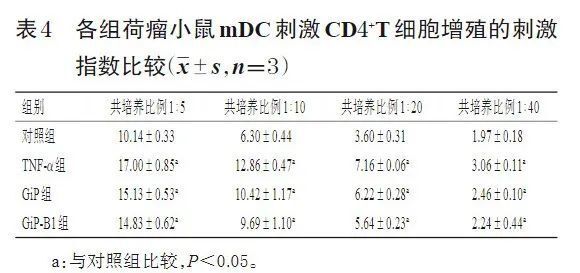
与对照组比较,TNF-α组、GiP组及GiP-B1组CD4-CTL上清液中IL-12p70(TNF-α组除外)、IFN-γ水平均显著升高,IL-4(TNF-α组除外)、IL-10(GiP组除外)水平均显著降低(P<0.05)。结果见表5。
效靶比分别为10∶1、20∶1、50∶1时,与对照组比较,TNF-α组(效靶比20∶1除外)、GiP组及GiP-B1组CD4-CTL对H22细胞的杀伤活性均显著增强(P<0.05)。结果见表6。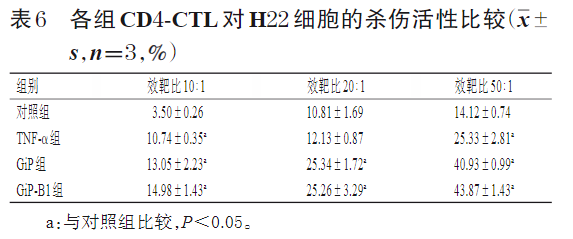
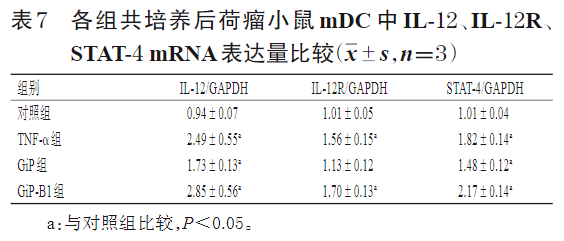
3.2.4 共培养后荷瘤小鼠mDC中IL-12、IL-12R、STAT-4 mRNA的表达情况 与对照组比较,TNF-α组、GiP组及GiP-B1组共培养后荷瘤小鼠mDC中IL-12、IL-12R(GiP组除外)、STAT-4 mRNA表达量均显著升高(P<0.05)。结果见表7。
3.2.5 共培养后荷瘤小鼠mDC中IL-12/NF-κB/STAT-4信号通路相关蛋白的表达情况 与对照组比较,TNF-α组、GiP组及GiP-B1组共培养后荷瘤小鼠mDC中IL-12Rβ2蛋白表达量和NF-κB p65、STAT-4蛋白磷酸化水平均显著升高(P<0.05)。结果见表8、图2。
4 讨论
多糖是一类具有调节免疫、抗氧化、抗肿瘤、抗炎等多种生物活性,且疗效良好、毒副作用低的天然高分子聚合物,在预防和治疗肿瘤中有广泛的应用前景。Wei等发现,铁皮石斛均一多糖DOPA-1能剂量依赖性地抑制肝癌细胞HepG2的生长;刘亚楠等发现,枸杞多糖能激活巨噬细胞RAW264.7并增强IL-6、IL-12分泌,发挥免疫调节作用。巨噬细胞的抗原提呈能力远远不如DC,并且DC迁移到淋巴结的能力对于启动T细胞介导的抗肿瘤反应是必需的。研究发现,黄芪多糖、三七多糖、人参多糖均能诱导DC成熟,从而激活T细胞,增强DC与T细胞的相互作用。因此,天然多糖通过促进DC成熟,提高肿瘤免疫疗效,在肿瘤治疗中具有一定的潜力。
表面标志物CD11c代表DC纯度,CD80与CD86是DC激活T细胞的共刺激分子,MHC-Ⅱ是DC抗原提呈分子,上述指标的阳性表达率升高表示DC的抗原提呈功能增强;DC分泌细胞因子的水平是其免疫功能是否成熟的重要指标之一,IL-12p70与IL-4是mDC的主要分泌细胞因子。本研究结果显示,经100 μg/mL GiP和100 μg/mL GiP-B1干预24 h后,荷瘤小鼠mDC的细胞活力、MHC-Ⅱ阳性表达率以及IL-12p70、IL-4水平均显著升高,CD11c、CD80、CD86阳性表达率也有升高趋势,提示GiP与GiP-B1对imDC有促成熟作用。
H22细胞因具备高成瘤性、遗传特性稳定、生长周期短、肿瘤位置与大小易于控制等优点,而成为抗肝癌作用的常用模型。H22细胞在培养过程中易于操作、培养条件易于控制,其所需培养基成分与本研究DC培养基大致相同。因此,本研究采用肝癌细胞H22评价GiP及GiP-B1诱导的mDC的抗肿瘤效应。DC能将肿瘤抗原交叉递呈给CD4+T及CD8+T细胞:前者极化为如抗肿瘤的T辅助性细胞1、促肿瘤的T辅助性细胞2等不同功能亚群;后者激活生成CTL细胞,发挥抗肿瘤效应。本研究将GiP与GiP-B1诱导的荷瘤小鼠mDC与CD4+T细胞共培养,以评估荷瘤小鼠mDC对CD4+T细胞的增殖能力和抗肿瘤效应。本研究结果显示,GiP及GiP-B1促成熟的荷瘤小鼠mDC均能显著刺激CD4+T细胞增殖,增强对H22细胞的杀伤活性。
为了阐明GiP及GiP-B1如何改善并增强mDC激活T细胞,进而发挥抗肿瘤的作用,本研究采用Transwell嵌套装置使mDC与CD4+T细胞间接共培养,以此维持上述细胞的功能和活力,同时只收集mDC用于检测分子水平的变化。结果显示,GiP及GiP-B1共培养后,荷瘤小鼠mDC中IL-12、IL-12R(GiP组除外)、STAT-4 mRNA表达量和IL-12Rβ2蛋白表达量以及NF-κB p65、STAT-4蛋白磷酸化水平均显著升高。这提示GiP及GiP-B1可能通过激活IL-12/NF-κB/STAT-4信号通路来提高荷瘤小鼠imDC、mDC免疫功能及其启动抗肿瘤免疫应答的能力。
综上所述,GiP及GiP-B1对荷瘤小鼠DC的成熟有较好的促进作用,能有效刺激CD4+T细胞增殖并增强CD4-CTL的抗肿瘤活性,其作用机制可能与激活IL-12/NF-κB/STAT-4信号通路有关。然而,GiP及GiP-B1基于调节DC功能的抗肿瘤作用机制有待进一步研究,二者抗肿瘤效应的差异也未做比较,今后还需进行系统的构效关系研究。
基金项目:国家自然科学基金项目(No.81960702);新疆维吾尔自治区重点实验室项目(No.XJDX1713)