
哺乳动物的海马体在情景记忆的形成与提取中扮演着至关重要的角色,特别是其亚区CA3在此过程中尤为重要。大量实验研究表明,海马体CA3区域的损伤会显著削弱动物在复杂情境中的记忆表现。此外,与空间记忆密切相关的位置细胞(place cell)序列活动,在解剖学层面最初起源于CA3。CA3的一个显著特征在于其神经元之间具有高密度的循环连接(recurrent connection),这正是吸引子网络(attractor network)的核心特点。吸引子网络能够从有限的线索中重构完整的记忆(pattern completion),还能够区分相似的线索,生成不同的输出(pattern separation)。这种能力对于在不同情境下准确提取特定记忆至关重要。因此,长久以来,人们推测海马体的记忆功能可能是通过吸引子网络实现的。尽管已有许多理论假设阐述了CA3可能通过哪些学习规则(learning rule)和神经通路实现这些功能,但直接的在体实验证据仍然匮乏。
2024年10月24日,贝勒医学院(Baylor College of Medicine)Jeffrey C. Magee和珍妮莉亚研究园( Janelia Research Campus)Sandro Romani合作(第一作者为李一丁)在Cell上发表了文章Mechanisms of memory-supporting neuronal dynamics in hippocampal area CA3。该研究旨在回答以下几个关键问题:1)何种形式的可塑性(plasticity);2)作用在哪些突触上使CA3能够编码位置信息;3)哪些输入通路(如有)能够更新吸引子网络中的活动状态;以及4)如此构建的吸引子网络在理论上具备哪些潜在能力。

研究人员训练头部固定的小鼠在跑步机上探索一维迷宫以获得水奖励,同时利用全细胞记录(whole-cell recording)在体监测CA3椎体神经元的膜电位变化。小鼠在熟悉任务后,约1/4 的CA3椎体神经元表现出位置细胞特性,即在特定位置发放阈上动作电位。研究人员观察到,CA3位置细胞的密度、神经元群体动作电位的频率、阈下膜电位的幅度以及θ频率震荡的强度,在整个跑道上呈现均匀分布。这恰好与吸引子网络的特性相吻合,即在网络中存在稳定的“能量”分布。在小鼠学习过程中,新的位置细胞不断涌现,那么这些位置细胞形成的机制是什么?研究人员在某些原本处于静默状态的细胞中观察到了一种独特的、持久的树突平台电位(dendritic plateau potentials),这一平台电位具有将静默细胞迅速转变为活跃位置细胞的能力。令人瞩目的是,这一平台电位可以在极短的时间内(下一圈行为中)显著改变神经元的膜电位,并且这一变化能够持续至实验结束。为了更细致地研究这一现象,研究人员在静默细胞中人为诱导树突平台电位。实验显示,绝大多数静默细胞(60/70)在诱导后成功转变为位置细胞。这一转变过程中膜电位的变化具有以下两个显著特点:1)对称性和双向性:平台电位±2秒窗口内,膜电位显著增强(potentiation),而在±2秒窗口外的则表现出减弱(depression);2)与突触强度负相关:膜电位变化的幅度与平台电位前的膜电位水平呈高度负相关。也就是说,先前较弱的突触增强得更多,而较强的突触则增强得较少。基于这些结果,研究人员认为这是一种新型的突触可塑性规则,与传统的Hebbian可塑性存在显著差异。研究人员将其命名为“行为尺度的突触可塑性”(Behavioral Timescale Synaptic Plasticity, BTSP),其主要特征包括:
1. 依赖于树突平台电位的触发;
2. 具有秒级的时间窗口,而非传统的毫秒级;
3. 可塑性表达非常快,可以通过一次学习(one-shot learning)快速实现;
4. 可塑性是对称的、双向的,同时包含突触增强和减弱。为了验证CA3位置细胞的产生是通过BTSP而非传统的Hebbian可塑性,研究人员在静默细胞中人为诱导了一串100Hz的高频动作电位,结果并未观察到任何显著的膜电位变化或位置细胞的转变。接下来,研究人员结合光遗传学技术,揭示了BTSP主要修改了CA3-CA3的循环连接的突触权重,而非齿状回(dentate gyrus,DG)投射到CA3的突触。由于BTSP的对称性,故在调整CA3-CA3突触权重时,可以产生方向上无偏的正反馈,从而构建出稳定的吸引子网络。CA3不仅接收来DG和自身的输入,还接收来自内嗅皮层(entorhinal cortex,EC)的输入。研究人员发现,EC的输入的功能在于动态更新吸引子网络中的活动状态。具体而言,当小鼠移动时,CA3“相邻”的位置细胞会依次激活,即小鼠的位置信息可以引导位置细胞的活动状态更新;然而,一旦阻断EC的输入,位置细胞的活动状态便无法更新,而是保持在阻断前的状态。由此,研究人员推测EC的输入类似于吸引子核心网络外的指导信号,指引吸引子网络中的活动峰(activity bump)向某个方向移动。上述两个实验结果暗示CA3位置细胞的产生机制并非通过直接整合来自EC的空间信息,而是通过调整与其他CA3位置细胞的突触连接权重实现的。EC作为一种外部输入,发挥了根据动物行为来更新CA3吸引子网络活动状态的作用。基于上述实验结果,研究人员构建了一个吸引子网络模型。该模型采用对称的BTSP规则调整核心网络中的连接权重,并通过外部输入来引导网络中活动峰的移动。这一模型很好地重复了研究人员在实验中观察到的现象。此外,相较于传统的Hebbian规则,基于BTSP规则构建的网络展现出更强的记忆存储能力。原因在于,BTSP规则包含靶向性突触权重减弱机制,而非随机减弱,这使得网络能够更好地对相似的输入进行去相关化处理,进而降低新记忆对已有记忆的干扰效应。综上,通过实验、理论分析和建模,该研究证实了海马CA3表现出吸引子网络的特性,并揭示了有效介导这些活动的细胞和环路机制。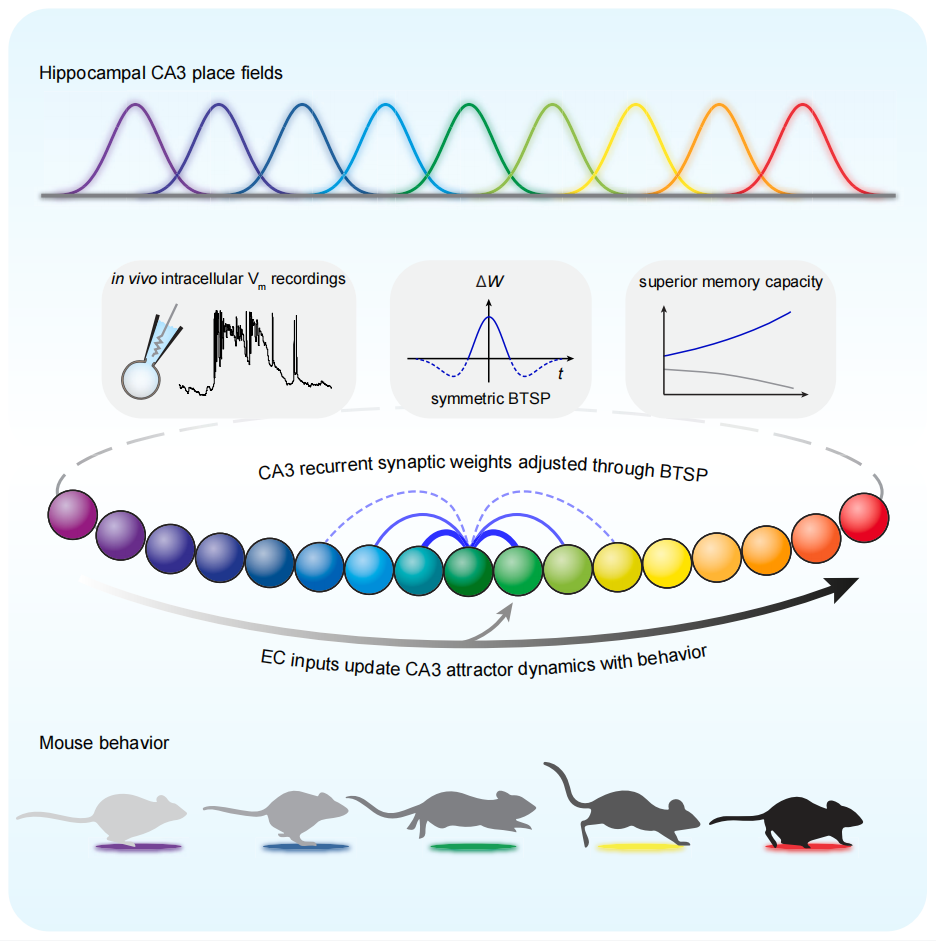
https://doi.org/10.1016/j.cell.2024.09.041